Dasypus bellus was about two to two and a half times the size of the living nine-banded armadillo (Dasypus novemcinctus).
Their diet was probably similar to the modern species, largely invertebrate animals, but the larger size of the fossil species may have allowed successful capture of small vertebrate animals, such as lizards and ground birds.
Source of Species Name: from the Latin word bellus, meaning “beautiful” or “pretty”. This is not one of the adjectives usually applied to armadillos, so Simpson was perhaps being ironic in this choice of name; but as he did not state the reason for the name in the original description, we will never know his extact intent.
Classification: Mammalia, Eutheria, Xenarthra, Cingulata, Dasypodoidea, Dasypodidae, Dasypodinae
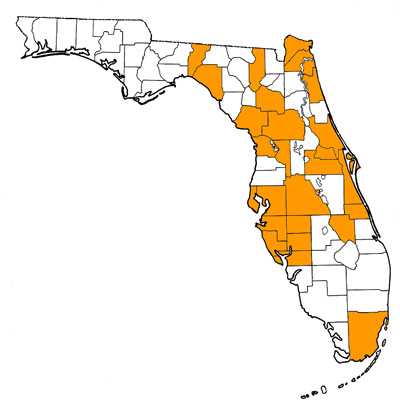
Figure 1. Map of Florida, with highlights indicating counties where fossils of this species have been found.
Overall Geographic Range
Southeastern, south-central, and midwestern United States, including occurrences throughout Florida (Fig. 1) and also in Georgia, South Carolina, Tennessee, West Virginia, Indiana, Iowa, Missouri, Nebraska, Kansas, and Texas (Klippel and Parmalee, 1984; Voorhies, 1987; Shubert and Graham, 2000). The type locality is Seminole Field, Pinellas County, Florida.
Indian River County—Luther Locality, Winter Beach; Vero Canal Site
Jefferson County—Aucilla River 2P
Levy County—McLeod Limerock Mine; Waccasassa River 1; Waccasassa River 2; Waccasassa River 3; Waccasassa River 4; Waccasassa River 5A; Waccasassa River 6; Waccasassa River 7; Waccasassa River 9A
Manatee County—Bradenton Field; Bradenton 51st Street
Marion County—Eichleberger Cave; Florida Lime Company Mine 2; Kendrick 1A; Mefford Cave 1A; Oklawaha River 1; Oklawaha River 3; Orange Lake 2A; Reddick 1A; Reddick 1B; Reddick 2C; Withlacoochee River 1A
On average, the osteoderms of the shell and the limb bones of Dasypus bellusare about two to two and a half times the size of those of the living nine-banded armadillo Dasypus novemcinctus. Fossils of Dasypus bellus have been found at many types of sites in Florida, including caves, sinkholes, river sites, coastal, and lake deposits. Isolated osteoderms are the most frequent type of fossil found, as is typical with cingulates generally. The two most common types of osteoderms recovered are the hexagonal elements that comprise most of the shoulder (pectoral) and pelvic regions of the carapace, the so-called buckler or immovable osteoderms (Fig. 2A), and the elongate rectangular elements from the movable bands (Fig. 2B), the imbricating or movable osteoderms (Holmes and Simpson, 1931; Hill, 2006). A partial articulated shell (Fig. 3) appears to have only seven or eight movable bands, although detailed study of this crushed specimen is needed to confirm this.
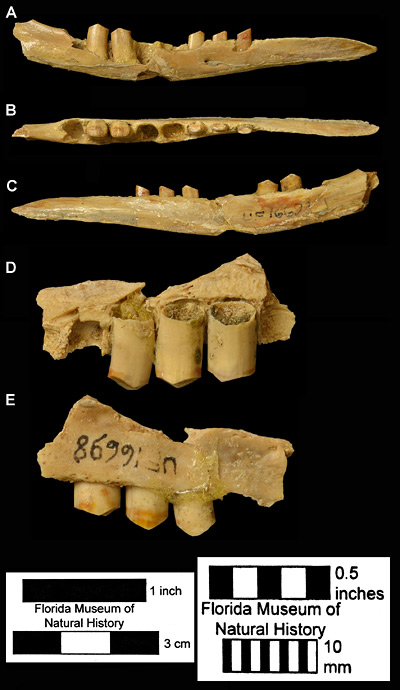
Like Dasypus novemcinctus, Dasypus bellus has small, simple, peg-like teeth (Fig. 4). The more posterior teeth have two flat wear surfaces that meet at a shallow angle, while the more anterior teeth have a single, slanted wear surface. The diet is thought to be similar to the modern species, largely invertebrate animals, but the larger size of the fossil species may have allowed successful capture of small vertebrate animals, such as lizards and ground birds. The skull is fragile and rarely preserves intact. The specimen shown in Figure 5 is the rare exception.
Figure 2. Osteoderms of Dasypus bellus. A, UF 136057, an immovable or buckler osteoderm in dorsal, lateral, and ventral views (top to bottom). Specimen from Haile 16A, Alachua County, Florida; early Pleistocene. B, UF/TRO 6831, a movable or imbricating osteoderm in dorsal, lateral, and ventral views (left to right). Specimen from Inglis 1A, Citrus County, Florida; early Pleistocene.
Figure 3. UF 61906, partial articulated carapace of Dasypus bellus from the Shell Materials Pit, Hillsborough County, Florida; early Pleistocene. This is the only known large section of a carapace of this species found articulated in Florida. The top image shows the entire specimen in dorsal view, while below is a close-up view of the pelvic buckler region.
Figure 4. UF 16698, associated right mandible and partial maxilla of Dasypus bellus from Haile 15A, Alachua County, Florida. A, lateral; B, dorsal; and C, medial (lingual) views of the mandible. D, medial (lingual); and E, lateral (buccal) views of the partial maxilla. Note the short, peg-like teeth. Left scale bar is for the mandible (A-C), while the right scale bar is for the maxilla.
Figure 5. UF 201289, nearly complete skull of Dasypus bellus from Inglis 2A, Citrus County, Florida; early Pleistocene. Above, dorsal view; below, ventral view. This is the most complete skull known for the species.
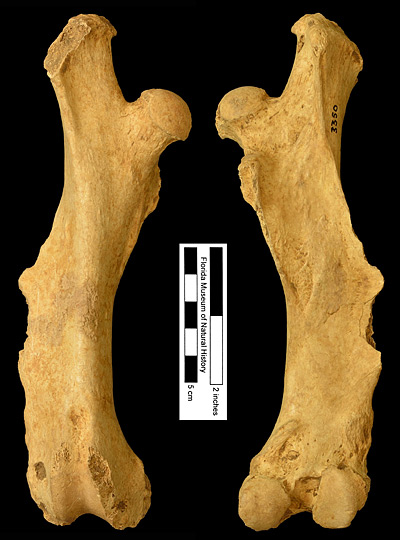
Figure 6. UF 3350, right femur of Dasypus bellus from Haile 8A, Alachua County, Florida; late Pleistocene. Left, anterior view; right, posterior view.
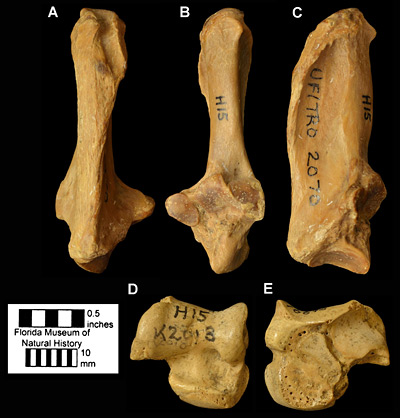
Figure 7. Ankle bones of Dasypus bellus from Haile 15A, Alachua County, Florida; early Pleistocene. A, posterior; B, anterior; and C, lateral views of UF/TRO 2070, right calcaneum. D, anterior; and E, posterior views of UF 16695, right astragalus.
Dasypus bellus is not especially closely related (or ancestral) to Dasypus novemcinctus, but instead likely shares a closer common ancestry with several species of large armadillos from the Pleistocene of South America, including Propraopus sulcatus and Propraopus grandis. This resemblance was noted in Simpson’s original description of the species in 1929, and Dasypus bellus is frequently referred to the genusPropraopus by South American authors (e.g., Pitana and Ribeiro, 2007). However, Rincón et al. (2008) found that features of the osteoderms shared by Dasypus bellus and Propraopus are also present in the largest living species of Dasypus, Dasypus kappleri, from tropical South America. Dasypus bellus and Dasypus kappleri also share a large, unreduced fifth digit on the manus, by which they differ from Dasypus novemcinctus that has a reduced manual fifth digit. As noted by Rincón et al. (2008), Dasypus kappleri is the type species of a generic name (Hyperoambon) that has priority over the genus Propraopus. In a more recent analysis, Castro et al. (2013) disputed a close relationship between Dasypus kappleri and Propraopus, but their study did not include Dasypus bellus.
There is a modest trend for increase in average size from the late Pliocene through the middle Pleistocene for Dasypus bellus (McDonald, 2005), although much less than in some other contemporaneous genera of xenarthrans (notably Holmesina, Megalonyx, and Paramylodon). Because of the apparent small amount of evolutionary change in Dasypus bellus, it is not a valuable species for biochronology.
Sources
Original Author(s): Richard C. Hulbert Jr.
Original Completion Date: September 12, 2013
Editor(s) Name(s): Richard C. Hulbert Jr.
Last Updated On: March 11, 2015
Auffenberg, W. 1957. A note on an unusually complete specimen of Dasypus bellus(Simpson) from Florida. Quarterly Journal Florida Academy of Sciences, 20(4):233-237.
Klippel, W. E., and P. W. Parmalee. 1984. Armadillos in North American late Pleistocene contexts. Pp. 149-160 in H. H. Genoways and M. R. Dawson (eds.), Contributions in Quaternary Vertebrate Paleontology: a Volume in Memorial to John E. Guilday. Carnegie Museum of Natural History, Special Publication No. 8.
McDonald, H. G. 2005. Paleoecology of extinct xenarthrans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45(4):313-333. (Download PDF)
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.