Xenosmilus hodsonae
Quick Facts
Common Name: Cookie-cutter Cat
A lion-sized, sabertoothed cat known only from Florida.
A mounted skeleton is on permanent display at the Florida Museum of Natural History.
Age Range
- Early Pleistocene Epoch; late Blancan to early Irvingtonian land mammal ages
- About 2.5 to 1.5 million years ago
Scientific Name and Classification
Xenosmilus hodsonae Martin et al., 2001
Source of Species Name: In honor of Debra Hodson, the wife of one of the men doing research on the species.
Classification: Mammalia, Eutheria, Laurasiatheria, Carnivora, Feliformia, Aeluroidea, Felidae, Machairodontinae, Homotherini
Alternate Scientific Names: Homotherium sp. of Berta (1995)
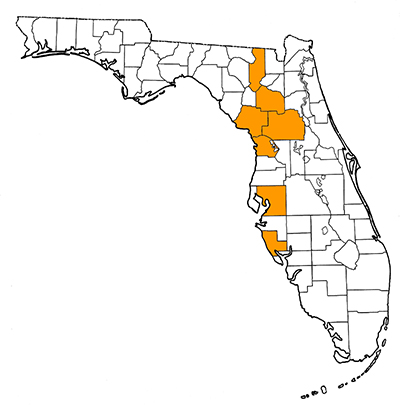
Overall Geographic Range
Xenosmilus hodsonae is only known from Florida. The type locality is Haile 21A in western Alachua County, Florida.
Florida Fossil Occurrences
Florida fossil sites with Xenosmilus hodsonae:
- Alachua County—Haile 7G; Haile 16A; Haile 21A.
- Citrus—Inglis 1A
- Columbia—Santa Fe River
- Hillsborough—Leisey Shell Pit 1A
- Levy— Waccasassa River 9A
- Marion—Withlacoochee River 1A
- Sarasota—Bass Point Waterway (identification tentative)
Discussion
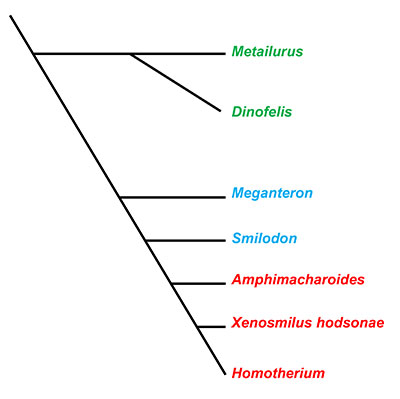
All true cats, extant and extinct, are classified in the mammalian family Felidae. All extant cats belong to the subfamily Felinae, which are characterized by the possession of conical canine teeth. In contrast, the Machairodontinae is an extinct subfamily of Felidae that consisted of cats with flatter, more knife-like like canines—commonly called the true saber-toothed cats. Machairodontinae has traditionally been divided into three tribes based largely on their anatomical similarities and inferred prey-capture strategies: the Smilodontini, Homotherini, and Metailurini. While Metailurini consists of species that are not easily classified into the other two tribes and likely represent an unresolved group at the base of the evolution of sabertoothed cats (Figure 2; Christiensen, 2013), species grouped in the tribe Smilodontini are often referred to as “dirk-toothed” cats, and those in the tribe Homotherini are often referred to as the “scimitar-toothed” cats. The dirk-toothed cats exhibited extended, bladelike canines and had stocky, powerfully built postcrania, suggesting that they were ambush predators. The scimitar-toothed cats possessed wide and short canines with long, agile legs for running down prey (Martin, 1980; Martin et al, 2011).
Xenosmilus hodsonae is a relatively recently described species of sabertoothed cat primarily known from two incomplete skeletons found in 1983 at Haile 21A in Alachua County, Florida. They were originally recovered by commercial fossil collectors. The less complete skeletons (UF 60000) was donated to the Florida Museum of Natural History, and the other (BIOPSI 101, the holotype of the species) was later obtained by Museum of Geology at Arizona State University. A mounted skeleton on display at the Florida Museum of Natural History is composed primarily of casts of the skull and limbs of BIOPSI 101, with those of UF 60000 used for bones missing from BIOPSI 101. When they were initially recovered, there was little interest in studying these two specimens because they were assumed to be those of Homotherium serum, a species of scimitar-toothed cat in the tribe Machairodontini, that had recently been described in literature (Rawn-Schatzinger, 1992). However, it was eventually noticed that these specimens possessed relatively short limbs for a scimitar-toothed cat, prompting a more detailed study (Martin et. al, 2011). In fact, Xenosmilus hodsonae contrasts from the traditional dichotomy of saber-toothed felids by combing the short canines of scimitar-toothed cats with the short-limbed, robust skeleton of a dirk-toothed cat (Martin et al., 2000). While a cladistic analysis of the cranial and dental characteristics has supported the hypothesis that the genera Xenosmilus and Homotherium are sister taxa, or more closely related to each other than to any other group (Figure 2; Christiansen, 2013), many features of the postcrania of Xenosmilus hodsonae resembles those of Smilodon, a smilodontine dirk-toothed cat (Martin et al., 2011).
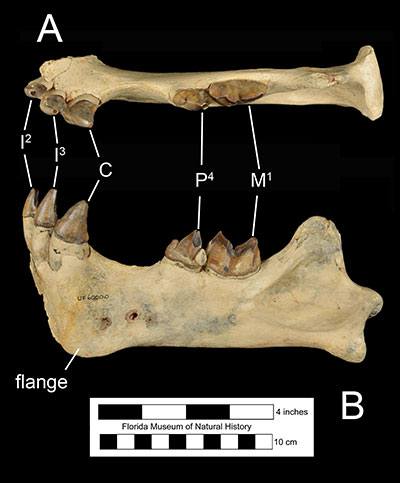
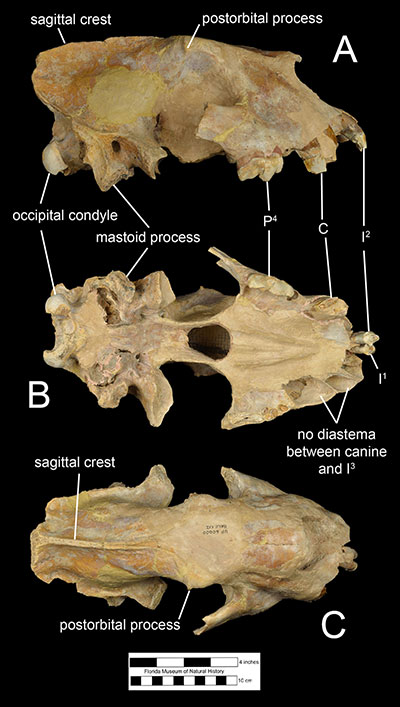
The general plan of the skull of Xenosmilus hodsonae is narrow and small in comparison to body size. The size of the skull is about as big as that of a small lion (Panthera leo) or a small individual of Smilodon fatalis, although its body size closer to the size of an average lion (Martin et al., 2011). The muzzle is shorter than in the genera Panthera and Smilodon with considerably narrow frontals and zygomatic arches displaying limited lateral flare (Figure 3). The cranium of Xenosmilus hodsonae is also characterized by a diminuitive postorbital process, large occipital condyles, and a relatively large mastoid process compared to the genus Homotherium. In addition, the mandibles of Xenosmilus hodsonae (Figure 4) have a smaller flange compared to Homotherium.Dentally, Xenosmilus hodsonae shares several characters with Homotherium which indicates that they are closely related
(Christiansen, 2013). Like other scimitar-toothed cats, the upper canines of Xenosmilus hodsonae were smaller and shorter than those of dirk-toothed cats; in fact, they were relatively shorter than even Homotherium. In addition, like other scimitar-toothed homotheres, all of the incisors of Xenosmilus hodsonae were relatively large compared to smilodontine cats, had serrated edges, and were positioned in a semicircular dental arcade. However, the incisor arcade of Xenosmilus hodsonae protruded more anteriorly than a typical scimitar-toothed cat (Martin et al., 2000; Martin et al., 2011). Unlike other scimitar-toothed cats, the sagittal crest of the skull of Xenosmilus hodsonae was relatively longer and more robust, similar to those of dirk-toothed cats. This suggests that the temporal muscle, which is used to close the mandible and generate bite force, was relatively larger and stronger than those of a typical scimitar-toothed cat. In addition, the elongate anatomy of the sagittal crest suggests that the temporal muscle exerted the greatest force along the anterior teeth (incisors and canines), unlike other saber-toothed cats, which experienced strongest bite forces along the carnassial teeth (Martin, 2000). This is particularly interesting as Xenosmilus hodsonae is unique amongst saber-toothed cats in lacking a diastema between its upper third incisors and upper canines, which effectively renders the anterior dental row a continuous, semi-circular cutting edge (figure 3).
This unique set of dental and cranial features prompted Martin et al. (2000 and 2011) to suggest that Xenosmilus hodsonae represented an example of a sabertoothed cat with a third type of feeding mode. As the common name “cookie-cutter cat” suggests, it is thought that Xenosmilus hodsonae used its entire anterior dental row to gouge large, semi-circular portions of flesh from its prey. This differs from dirk-toothed cats, which are believed to have killed by slicing prey with their larger canines and removing smaller portions of flesh, and from scimitar-toothed cats, which are believed to have removed strips of flesh from their prey (Martin et al., 2001). The animal’s large, uniquely oriented temporal muscles are thought to have helped the cookie-cutter cat exert its killing bite.
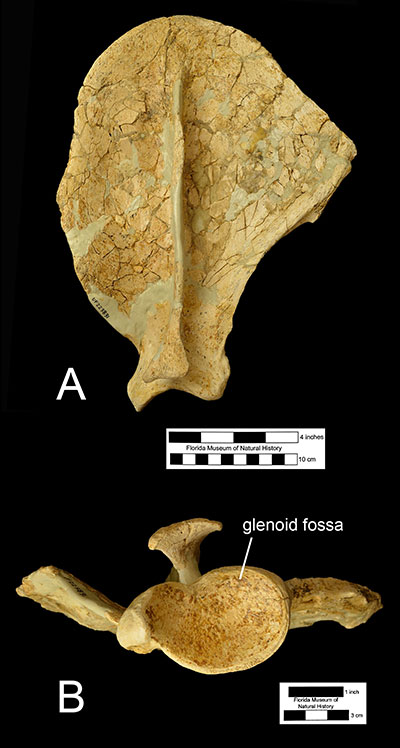
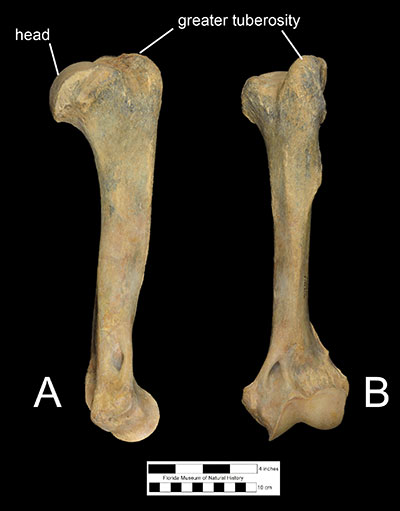
Features of the postcranial elements suggest a powerfully built ambush predator capable of grappling with large prey. The scapula (Figure 5) is larger than that of Smilodon and has a hefty, oval-shaped glenoid fossa, suggesting that the shoulder was capable of moving the forelimbs laterally while wrestling prey. The humerus (Figure 6) is narrower than that of Smilodon but possesses some features that are more robust including a larger and more rounded humeral head, and a taller, broader greater humeral tuberosity. Corresponding muscle attachments indicate a wider range of movement in the shoulder joint. The ulna is shorter than the humerus and shorter than in Smilodon. Features on the articular surfaces of humerus, radius, and ulna suggest that Xenosmilus hodsonae was capable of pronating the forearm to a larger degree than other large cats, which would have facilitated its ability to grasp struggling prey and perhaps even climb trees. The radius and hand bones are also curved in such a way to suggest that the animal could exert great force to pull in its prey closer (Martin et al., 2011).
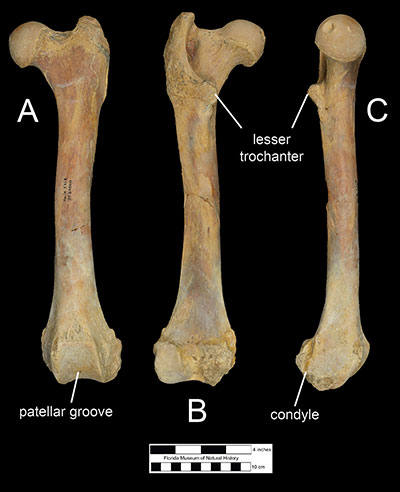
The hindlimbs of Xenosmilus hodsonae also show several specializations for prey capture. The femur (Figure 7) is relatively shorter and more robust than even those of bears or giant panda. Compared to other large cats, the lesser trochanter of the femur was larger, suggesting that the psoas major muscle, which attaches there, was well developed. A more powerful psoas major muscle would have had kept the hips of the animal steady while it reared up on its hind legs while wrestling prey with its forelimbs. The femur also has a relatively wide patellar groove, which supported a relatively large patella (Figure 8). This indicates that Xenosmilus hodsonae had large quadriceps muscles which stabilized the knee during prey capture. However, the femoral condyles of Xenosmilus hodsonae protrude further posteriorly than other large cats, suggesting that this animal also had more flexible knees, which would have been useful while stooping down to ambush prey. Furthermore, features on the tibia and ankle bones suggest that Xenosmilus hodsonae held its foot in a plantigrade position, similar to bears (Martin et al., 2011).

Overall, Xenosmilus hodsonae was a specialized ambush predator that probably exhibited a prey capture style different from those of the scimitar-toothed and dirk-toothed saber-toothed cats. Xenosmilus hodsonae probably lived and hunted in savanna or woodland with sufficient vegetation to facilitate its unique style of ambush hunting. With its relatively small head, short, stocky limbs, and plantigrade posture, Xenosmilus hodsonae also probably looked more like a bear in body proportions than a cat (Martin et al., 2000; 2011). Since the discovery and description of the two skeletons from the Haile 21A locality, specimens of Xenosmilus hodsonae have been recognized from several other sites in Florida (these had previously identified as Homotherium [Webb, 1974; Berta, 1995]). No other records exist for the species outside of Florida. However, recent discoveries of specimens attributed to the genus Xenosmilus from Uruguay (Mones and Rinderknecht, 2004) and Arizona (White et al., 2005) provide hope that with the recovery of more fossils, the evolution of this unique sabertoothed genus may one day be better understood.
Sources
- Original Author: Arianna Harrington
- Original Completion Date: April 13, 2015
- Editor(s) Name(s): Arianna Harrington, Natali Valdes, Ricard C. Hulbert Jr.
- Last Updated On: April 27, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida