Neohipparion eurystyle
Quick Facts
Common Name: none
This moderate-sized hipparionine horse ranged from central Mexico to Nebraska and east to central Florida.
Neohipparion eurystyle and the closely related, larger species Neohipparion gidleyi (not known from Florida) represent the culmination of adaptation towards a pure grass diet (hypergrazing) among North American hipparion horses.
Age Range
- Late Miocene to earliest Pliocene epochs; Hemphillian 2, 3, and 4 land mammal ages
- About 7 to 4.5 million years ago
Scientific Name and Classification
Neohipparion eurystyle Cope, 1893
Source of Species Name: Not explicitly stated in Cope’s (1893:43-45) original description. The name was formed by combining the Greek words eurys (‘broad’ or ‘wide’) and stylos (‘pillar’ or ‘column’), and probably refers to the “relatively great width of the metaconid-metastylid column” in the species that Cope considered characteristic of the species.
Classification: Mammalia, Eutheria, Eutheria, Laurasiatheria, Perissodactyla, Hippomorpha, Equidae, Equinae, Hipparionini
Alternate Scientific Name: Equus eurystylus, Hipparion eurystyle, Hesperohipparion eurystyle, Hipparion phosphorum, Neohipparion phosphorum
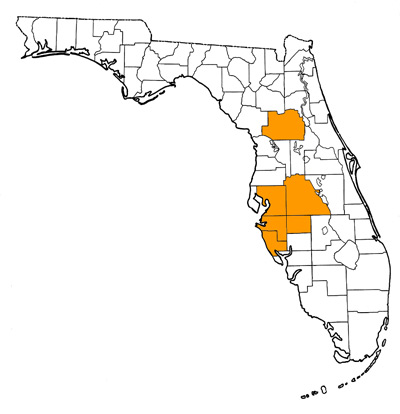
Overall Geographic Range
During the late early Hemphillian (Hh2), Neohipparion eurystyle is known only from the southeast Gulf Coastal Plain (Florida and Alabama; Hulbert and Whitmore, 2006). During the late Hemphillian (Hh3 and Hh4), its range expanded west to include all of Texas, north to Nebraska and possibly North Carolina, and south to central Mexico. Possibly known from California, but diagnostic material has yet to be found on the West Coast. The type locality is the Goodnight Beds, Palo Duro Canyon, Randall County, Texas (Cope, 1893; Dalquest, 1981).
Florida Fossil Occurrences
Florida fossil sites with Neohipparion eurystyle:
- Hardee County—Hardee Complex Mine (C.F. Industries); Fort Green Mine
- Hillsborough County—Big Four Mine; Four Corners Mine; Kingsford Mine; Leisey Shell Pit 1C
- Manatee County—Braden River Site; Fort Green Mine; Four Corners Mine; Manatee County Dam Site; Port Manatee Site; SR64 Site
- Marion County—Moss Acres Racetrack Site; Mutual Mine of Camp Phosphate Company; Withlacoochee River 4A Site; Withlacoochee River 4X Site; Withlacoochee River 5E Site
- Polk County—Brewster Mine; Chicora Mine; Fort Green Mine; Fort Meade Mine (Gardinier), including Whidden Creek Site; Hookers Prairie Mine; Kingsford Mine; Mineco Mine; Nichols Mine; North Palmetto Mine; Palmetto Mine; Payne Creek Mine; Phosphoria Mine; Silver City Mine; South Fort Mead Mine (Mosaic)
- Sarasota County—Lockwood Meadows Site
Discussion
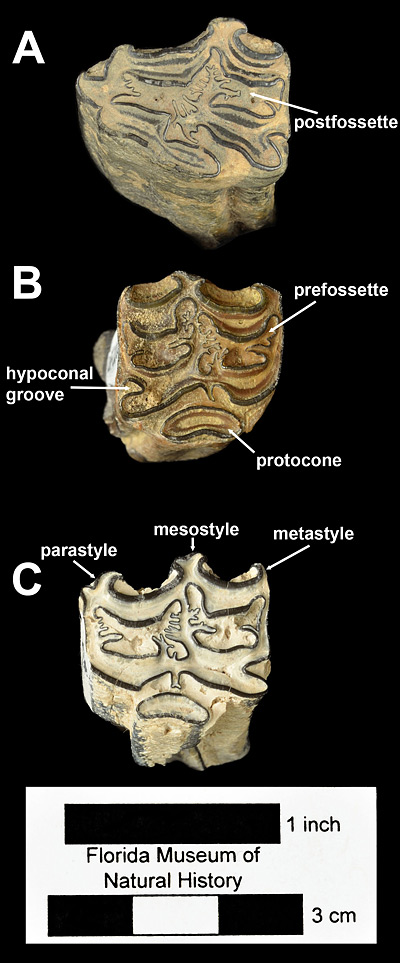
Many separate species names were originally proposed for this moderate-sized hipparionine horse. MacFadden (1984) synonymized Neohipparion phosphorum (Simpson), Neohipparion arrellanoi (Stirton), Neohipparion floresi (Stirton), Neohipparion otomii (Mooser), Neohipparion monias (Mooser), and Hesperohipparion stirtoni (Dalquest) with Neohipparion eurystyle (Cope), noting that the supposed differences in size and tooth characters between them are relatively trivial and more similar in scope to those found between populations of the same species in other equids. Neohipparion eurystyle and the closely related, larger species Neohipparion gidleyi (not known from Florida) represent the culmination of adaptation towards a pure grass diet (hypergrazing) within the genus Neohipparion (Rensberger et al., 1984; Hulbert, 1986). As they evolved their teeth for this specialized diet, many of the same features that were found in the teeth of other hypergrazing horses (including Equus) appeared independently. Cope mistakenly thought that the holotype specimen, a partial lower tooth, belonged in the genus Equus because of these similarities. Features shared by equid hypergrazers include extremely tall crowns, elongated protocones and metaconids-metastylids, and flattened margins of the protocone, protoconid, and hypoconid (Figs. 2-3). The limbs of Neohipparion eurystyle were long and slender, and retained small but complete lateral digits (Sondaar, 1968). Neohipparion eurystyle and Neohipparion gidleyi are the two youngest known species in the genus, both becoming extinct at the Hemphillian-Blancan boundary about 4.5 Ma.
Simpson (1930) based the species Neohipparion phosphorum on a single upper molar found in a phosphate mine near the town of Brewster, Florida. At the time, Neohipparion eurystyle was only known from lower teeth, so no direct comparison between them could be made. Since the publication of Simpson’s paper in 1930, over 300 additional specimens of this species have been found in the phosphate mines of Central Florida, mostly isolated teeth, both uppers and lowers. A few sets of associated teeth and partial jaws are known from the phosphate mines. Four relatively complete mandibles, two that include matching right and left sides, were recovered from the Moss Acres Racetrack Site in western Marion County (Fig. 4). Unfortunately, none of these were associated with skeletons or skulls, as was the case for some other species of horses at this site. The only known skulls for this species were collected in Mexico (MacFadden, 1984).
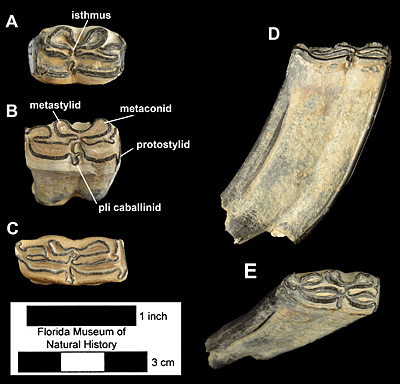
The adult cheekteeth (premolars and molars) of Neohipparion eurystyle are highly distinctive and relatively easily identified. If slightly worn and unbroken, they will be over 65 mm (2.5 inches) tall, more than any other North American horse with the exception of Nannippus peninsulatus and the various species of Astrohippus and Equus. The enamel pattern on the upper cheekteeth (Fig. 2) includes an elongate protocone that is isolated except on some second premolars (Fig. 2A) and that has a lingual border that is either concave, flat, or sinuous, not convex. Protocone length is very often greater than half that of the entire occlusal surface. The fossettes are moderately complicated, but not to the degree of Cormohipparion emsliei. Styles are very strong, with the metastyle curling anteriorly and the parastyle curling posteriorly and the distal end of the mesostyle expands laterally, forming a ‘T’-shape (Fig. 2). The hypoconal groove is broadly open to near the base of the crown. The lower cheekteeth are equally distinct (Figs. 3-4). The metaconid-metastylid are long and narrow; the isthmus is complete and not fully divided by the shallow ectoflexid, even in molars; the pli caballinid is very prominent, extending laterally farther than the outer margin of the hypoconid, and frequently is secondarily folded; and the protostylid is also prominent and curves posteriorly. The lower deciduous premolars lack ectostylids. In late Hemphillian faunas, compared to the teeth from contemporaneous horse species, those of Neohipparion eurystyle are larger than those of Pseudhipparion simpsoni, Nannippus aztecus, or Nanippus lenticularis; smaller than those of Dinohippus interpolatus or Dinohippus mexicanus; and about the same as those of Cormohipparion emsliei, Astrohippus ansae and Astrohippus stockii. In addition to more numerous fossette plications and shorter crowns, cheekteeth of Cormohipparion emsliei can be distinguished from those of Neohipparion eurystyle by shorter protocone; parastyle, metastyle, and protostylid not curved; mesostyle not ‘T’-shaped; deeper ectoflexids and smaller pli caballinid on lower molars, and a prominent ectostylids on deciduous lower premolars. The cheekteeth of Astrohippus have very different enamel patterns than those of Neohipparion eurystyle, with connected protocones, simple fossettes, rapid loss of hypoconal grooves, weak to absent metastyles, and lack of plis caballinid and protostylids. Among Florida equids, the closest species in terms of dental patterns is Neohipparion trampasense, which makes sense as it is the likely ancestor of Neohipparion eurystyle (Hulbert, 1987). The cheekteeth of Neohipparion trampasense are shorter than those of Neohipparion eurystyle (maximum about 60 mm). In uppers, the protocone is shorter, the mesostyles are wider and not distally expanded, and the parastyle and metastyle are not curved. In the lowers, the molars have deeper ectoflexids, especially in middle and late wear.

Sources
- Original Author(s): Richard C. Hulbert Jr.
- Original Completion Date: February 16, 2012
- Editor(s) Name(s): Richard C. Hulbert Jr.
- Last Updated On: March 2, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida