Alligator olseni
Quick Facts

Common Name: Olsen’s Alligator
Adult Alligator olseni specimens are generally smaller than adult Alligator mississippiensis specimens, with few individuals exceeding more than seven to eight feet in length, while fossils of Alligator mississippiensis from Florida frequently exceeded ten feet.
Although many different evolutionary hypotheses exist, Alligator olseni continues to be one of the best known extinct species within the genus and an important focus of research by crocodylian specialists.
Age Range
- Early Miocene; Hemingfordian land mammal age
- About 19 to 16 million years ago
- Possibly extends back to very early Miocene (late Arikareean), about 23-22 million years ago
Scientific Name and Classification
Alligator olseni White, 1942
Source of Species Name: Named for Russel Olsen, who prepared the original holotype specimen for the Museum of Comparative Zoology at Harvard University (White, 1942).
Classification: Reptilia, Archosauria, Crocodylomorpha, Crocodylia, Alligatoridae, Alligatorinae
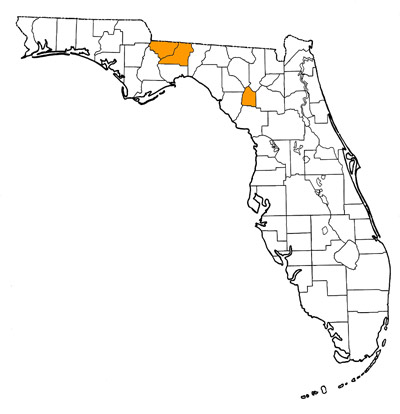
Overall Geographic Range
Southeastern United States, with occurrences in southeastern Texas (possibly) and Florida. The type locality is Thomas Farm, near Bell, Gilchrist County, Florida (White, 1942).
Florida Fossil Occurrences
- Gadsden County—Midway Local Fauna; Milwhite Gunnfarm Mine
- Gilchrist County—Thomas Farm
- Leon County—Griscom Plantation; Seaboard Local Fauna
Discussion
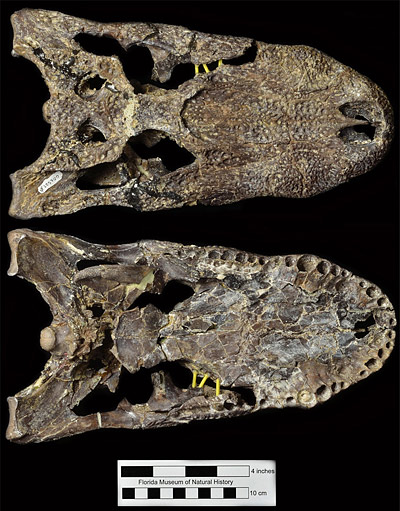
Alligator olseni is an intermediate species nested between primitive and derived members of the genus Alligator (Malone, 1979; Brochu, 1999, 2004, 2011; Snyder, 2001, 2007). It was initially described from Thomas Farm by White (1942), and all the more complete specimens representing this species have since been collected from the same locality (Brochu, 1999; Snyder, 2001, 2007). Excavations at Thomas Farm continue on a regular basis, and Alligator olseni fossils are still discovered and collected by palaeontologists and volunteers alike (Fig. 2).
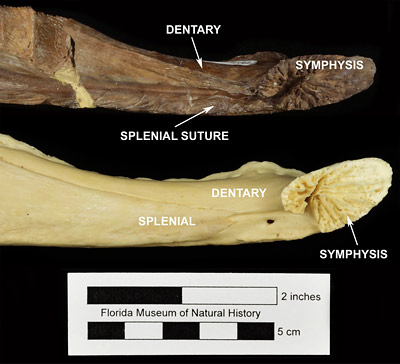
The skeleton of Alligator olseni looks very similar to that of the modern American alligator (Alligator mississippiensis), but they can be distinguished by some subtle characteristics. The splenial bone, found on the internal (medial) surface of the lower jaw in crocodylians, rarely preserves as a fossil. However, the preserved suture lines on the dentary bone are typically visible and can be used to determine how far anteriorly the splenial extended. In Alligator olseni, the splenial usually extends into the mandibular symphysis (Fig. 3; White, 1942; Malone, 1979; Snyder, 2001, 2007). In Alligator mississippiensis, the splenial does not extend this far, and ends posterior to the mandibular symphysis (Fig. 3).
In addition, adult Alligator olseni specimens are generally smaller than adult Alligator mississippiensis specimens, with few individuals from Thomas Farm likely exceeding more than about seven to eight feet (2.5 meters) in length (Meylan et al., 2001). Fossils of Alligator mississippiensis from Florida frequently represent individuals that exceeded ten feet (3.3 meters) in length, and are found in much younger deposits (Meylan et al., 2001). The snout of Alligator olseni is also comparatively blunter than the snout of Alligator mississippiensis, and there is often a higher degree of ornamentation on the dorsal skull surface near the orbits in Olsen’s alligator than in the American alligator. For more on the osteology of Alligator olseni and comparisons with other named species of Alligator, see White (1942) and Snyder (2007).
There are some other transitional forms potentially separating Olsen’s alligator and the American alligator, although their exact placement within the genus Alligator is debated. Brochu’s (1999, 2004, 2011) analyses placed Alligator sinensis, the extant Chinese alligator, as a closer relative to the modern American alligator than Alligator olseni, but Snyder (2001, 2007) found it basal to Alligator olseni and Alligator mississippiensis. This debate brings into focus the question of how and when the ancestor of the Chinese alligator dispersed into Asia from North America. See Brochu (1999, 2003) and Snyder (2001, 2007) for more on this topic. The validity of Alligator thomsoni Mook, 1923 and Alligator mefferdi Mook, 1946, two other Miocene Alligator species, are also disputed. Malone (1979) considered both to be synonymous with Alligator mississippiensis, while Brochu (1999) and Snyder (2001, 2007) regarded both to be valid species closely related to the modern American alligator. The placement and validity of different species within the genus Alligator is an ongoing endeavor subject to frequent debate.
Other fossils from Florida assigned to Alligator olseni were collected from the Midway and Milwhite Gunnfarm Mine localities of Gadsden County, as well as the Griscom Plantation and Seaboard localities of Leon County (Auffenberg, 1957; Olsen, 1964, 1968; Stoutamire, 1975). This fossil material consists of isolated teeth, osteoderms, and a few fragmentary vertebrae and skull bones. These sites are estimated to be similar in age with, and geographically close to, the type locality (Fig. 1; Woodburne, 2004). In addition, the morphology and sizes of the fossils from these sites are consistent with similar elements from Thomas Farm (Auffenberg, 1957). Teeth and partial osteoderms of the unrelated crocodylian Gavialosuchus are also potentially present at some of these sites (Auffenberg, 1957). This complicates the assignment of the isolated remains, but the simplest way to differentiate between the teeth and osteoderms of the genus Alligator and the genus Gavialosuchus is to compare them with specimens of known affinity (Meylan et al., 2001). Osteoderms, the highly pitted dermal bones forming the scutes running along the dorsal surface (back) of crocodylians, are essentially flat and possess no midline dorsal keel in Gavialosuchus. However, in Alligator, the osteoderms possess a pronounced midline dorsal keel and are not flat (Fig. 4). Regarding dentition, the teeth of Gavialosuchus are typically narrower in diameter and taper to a sharper tip, while those of Alligator are usually blunter at the tip and wider in diameter (Fig. 5).


Albright (1994) reported Alligator olseni fossils from the Toledo Bend locality in Texas near its border with Louisiana, although its age differs from Thomas Farm. This locality is estimated at approximately 23-22 million years ago in the late Arikareean (very early Miocene), which is notably older than the Hemingfordian (early Miocene) age of Thomas Farm (Woodburne, 2004). Albright (1994) used the characteristic splenial to conclude that the material from Toledo Bend represents Alligator olseni, although this was not confirmed by Snyder (2007) or mentioned by Brochu (1999). A cast of an alligatorid tooth (Fig. 4C) from Toledo Bend in the FLMNH collections (the original fossil is housed in the LSU Museum of Geology) is much larger than definitive Alligator olseni specimens from Florida, and may instead represent a different, larger alligatorid taxon. Until the fossils described by Albright (1994) are reexamined, the assignment of the Toledo Bend material to Alligator olseni is questionable. However, if this material does represent Alligator olseni, Albright’s (1994) record would provide significant range extensions in body size as well as geographic and geologic occurrence for the species.
Since its initial discovery and description (White, 1942), Alligator olseni has continued to be an important taxon when attempting to determine evolutionary relationships within Alligator. The Thomas Farm locality regularly produces well-preserved specimens, including several skulls found by museum palaeontologists and volunteers over the past two decades. Although the evolutionary relationships of the species within the genus Alligator are not absolutely resolved and many different evolutionary hypotheses exist (Mook, 1946; Malone, 1979; Brochu, 1999, 2004, 2011; Snyder, 2001, 2007), Alligator olseni continues to be one of the best known extinct species within the genus and an important focus of research by crocodylian specialists.
Sources
- Original Author(s): Evan T. Whiting
- Original Completion Date: December 8, 2012
- Editor(s) Name(s): Richard C. Hulbert Jr., Natali Valdes
- Last Updated On: February 25, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida