Hemiauchenia macrocephala

Quick Facts
Common Name: large-headed llama
The most complete skeleton of this species was found near Lecanto in Citrus County, Florida. It is on permanent display at the Florida Museum of Natural History.
One of three species of Hemiauchenia that lived in Florida during the Blancan land mammal age, but the only one known to survive into the Irvingtonian. It persisted until the latest Pleistocene and would have been a contemporary with PaleoIndians in the state.
Modern studies reveal that it was a dietary generalist, with some individuals subsisting on a pure browsing diet, others on a nearly pure grazing diet, but a majority eating plants of both types.
Age Range
- Early to late Pleistocene Epoch; late Blancan to very late Rancholabrean land mammal ages
- About 2.6 million to 11,000 years ago
Scientific Name and Classification
Hemiauchenia macrocephala Cope, 1893
Source of Species Name: Not explicitly stated in Cope’s (1893) original description. The species name is formed by the combination of the Greek words makros meaning ‘large’ or ‘long’ and –kephalos meaning ‘head’. Cope’s intent here is not clear, as at the time he named the species he only had the lower jaw, and no fossils from the skull. He did state that the lower jaw was “considerably more elongate” than that of the living Lama lama, so the macro– portion of the specimen name may have referred to its great length rather than simply large size.
Classification: Mammalia, Eutheria, Laurasiatheria, Artiodactyla, Camelidamorpha, Tylopoda, Cameloidea. Camelidae, Camelinae, Lamini
Alternate Scientific Names: Holomeniscus macrocephalus; Tanupolama macrocephala; Lama stevensi; Lamahollomani; Camelus americanus
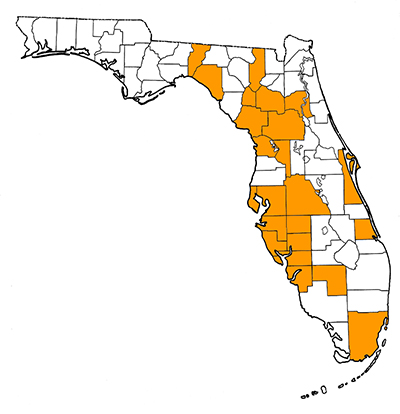
Overall Geographic Range
The only records of this species during the Blancan are from Florida. During the Irvingtonian and Rancholabrean, its range expanded westward and northward to include Mississippi, South Carolina, Nebraska, Kansas, Texas, Oklahoma, New Mexico, Idaho, Utah, and California. The northernmost record is from southeastern Alberta, Canada. It is also known from northern and central Mexico (Bravo-Cuevas et al., 2012). The type locality is Rock Creek, an Irvingtonian site in Briscoe County, Texas (Cope, 1893; Webb, 1974).
Florida Fossil Occurrences
Florida fossil sites with Hemiauchenia macrocephala:
Discussion
Hemiauchenia macrocephala is a member of the family Camelidae, which today includes the camels of the Old World (the tribe Camelini) and the llamas of South America [the tribe Lamini (note the single letter ‘l’ to start the scientific name, unlike the double ‘ll’ that starts the common name)]. The family Camelidae originated and diversified in North America (Honey et al., 1998), only to become extinct here and survive on other continents. At least four extinct subfamilies of the Camelidae are known from fossils in North America: the extinct Miolabinae, Protolabidinae, Stenomylinae, and Floridatragulinae, as well as the extant Camelinae. The two tribes within the Camelinae, Camelini and Lamini, originated during the middle Miocene in North America, with the former dispersing to Asia across the Bering Land Bridge and the latter dispersing to South America through the Panamanian Isthmus. Hemiauchenia macrocephala, like all other Pleistocene camelids from Florida, is a member of the Lamini (Figs. 2-3). These animals are more properly referred to as llamas than as camels when using common names, as they are more closely related to and more greatly resemble modern llamas than camels.
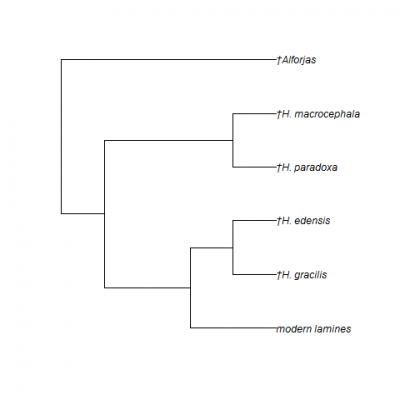
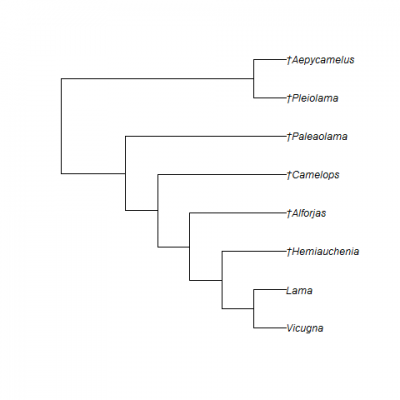
The genus Hemiauchenia was first named for several fossil species from South America with relatively long, slender limb bones and hypsodont molars (Webb, 1974). For almost a century, similar fossil llamas were recovered from North America, but were assigned to other genera, such as Holomeniscus, Lama, and Tanupolama. Webb (1974) was the first researcher to recognize the close relationship between South American Hemiauchenia and these North American species, placing them in the same genus. This conclusion has been accepted by all subsequent researchers (e.g., Breyer, 1977; Dalquest, 1992; Honey et al., 1998; Jiménez-Hidalgo and Carranza-Castañeda, 2010; Bravo-Cuevas et al., 2012; Scherer, 2013). However, the number of valid North American species of Hemiauchenia is still controversial, and if the results of Scherer (2013) are correct, the genus is paraphyletic and its species need to be divided among multiple genera (Figs. 2-3).
The species Holomeniscus macrocephalus was named by Cope in 1893 on the basis of a single mandible from the Texas Panhandle. Similar species were later named from Nebraska (Camelus americanus), Oklahoma (Lama hollomani), and California (Lama stevensi). Stock (1928) named the new genus Tanupolama for these species with Tanupolama stevensi as the type species of the genus. Hibbard and Dalquest (1962) synonymized Tanupolamastevensi and Tanupolama americana with Tanupolama macrocephala, reducing some of the clutter. Webb (1974) added Tanupolama hollomani to this synonymy, and transferred them to the genus Hemiauchenia. Dalquest (1992) went even further, synonymizing Hemiauchenia macrocephala with the larger Hemiauchenia blancoensis and Hemiauchenia seymourensis. Dalquest’s synonymy is being followed by the Paleobiology Database as of June 2015, but it has not generally been accepted by others working on camelids (e.g., Honey et al., 1998; Meachen, 2005; Bravo-Cuevas et al., 2012). The current practice of paleontologists at the Florida Museum of Natural History is to recognize Hemiauchenia macrocephala and Hemiauchenia blancoensis as distinct species.

The presence of two species of llamas in the late Pleistocene of Florida was first realized by Simpson (1929), which he referred to as Tanupolama mirifica and Tanupolama americana. Bader (1957) noted the same situation at the Arredondo site. The specimens they regarded as Tanupolama americana would now be called Hemiauchenia macrocephala. It seems that Webb (1974) was the first to actually use Cope’s species name macrocephala for specimens from Florida. The species is now known from many Florida localities (see list above and Fig. 1) ranging in age from late Blancan to latest Pleistocene. Very large samples are now known from the Leisey Shell Pits (confusingly described by Webb and Stehli, 1995 under the name Hemiauchenia seymourensis), as well as a nearly complete skeleton that is mounted on permanent display at the Florida Museum of Natural History. A catastrophic assemblage composed mostly of juvenile and subadult individuals of Hemiauchenia macrocephala was recovered from Leisey Shell Pit 3A during the winter of 1986-1987.
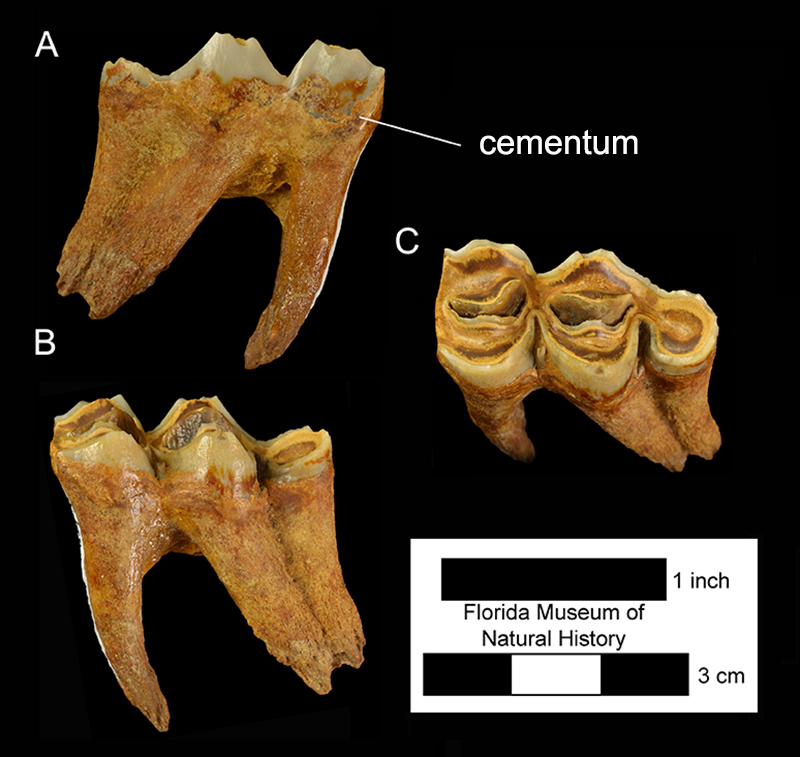
Although it has been partially restored for exhibition, the skull of UF 205750 from the Lecanto 2A locality in Citrus County is the most complete one known for the species (Fig. 4). It has yet to be studied in detail. There is a small, caniniform third incisor (I3) in the posterior portion of the premaxilla. A diastema separates the I3 from the larger, true upper canine on the anterior end of the maxilla. An equally long diastema separates the canine from a caniniform first premolar (P1). A diastema about twice as long separates the P1 from the P3 (the P2 is absent; Fig. 10). The P3 through third molar (M3) form a continuous series. The P3-M3 are moderately hypsodont and the outer enamel is coated with a thin layer of cementum. The inner or lingual crests of the molars tend to be more U-shaped than V-shaped (Fig. 5).
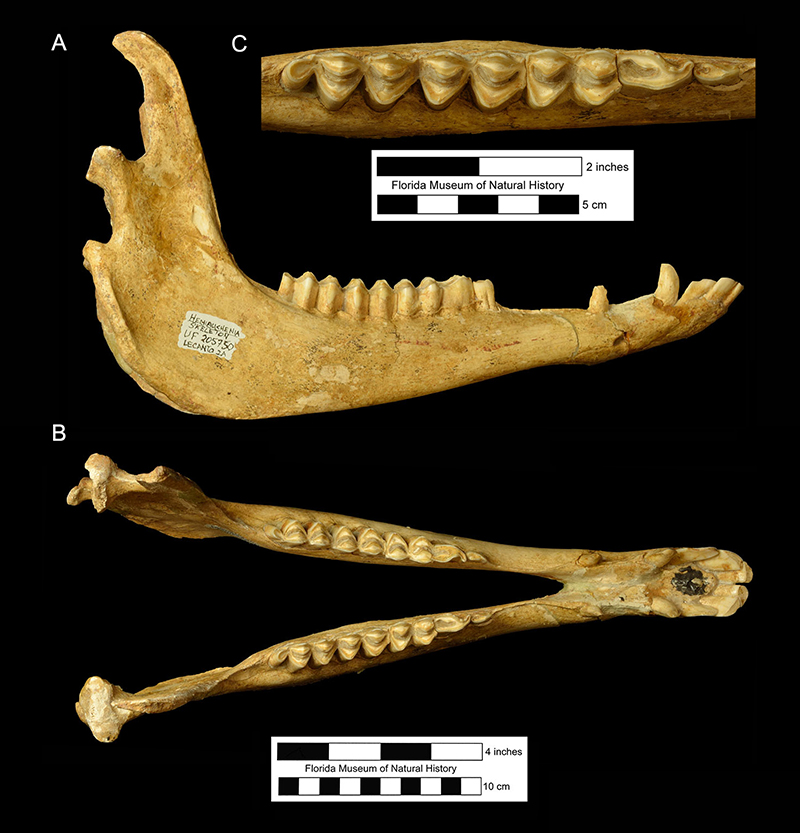
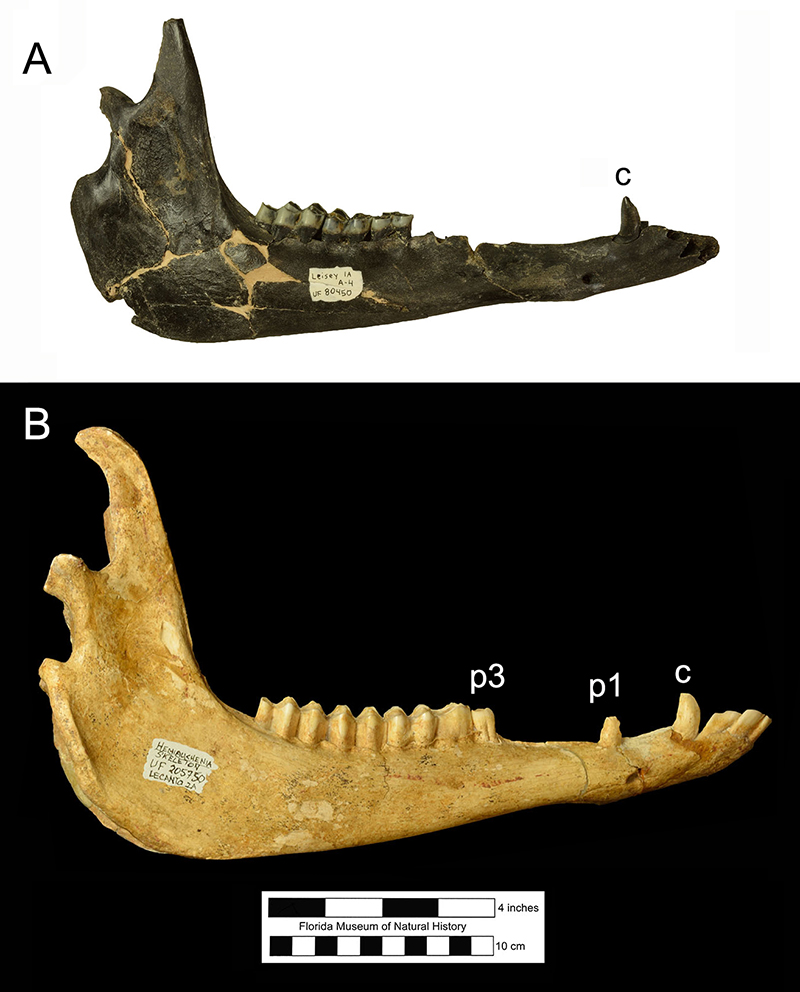
The mandible has a long rostral region and a deep ramus below the molars (Fig. 5). There are three large, procumbent incisors with long crowns, followed by the sharp canine. A second caniniform lower tooth, the first premolar (p1), is located just in front of or directly above the mental foramen. Two sizes of p1s can be observed in the Leisey Shell Pit 1A sample of adult mandibles, suggesting sexual dimorphism. The lower cheekteeth consist of a small p3, a larger p4, and three molars. Like the uppers, these teeth are hypsodont and have a cement coating (Fig. 8). The p4 is simple, lacking the outer groove and multiple inner folds found in the same tooth of Palaeolama mirifica (Fig. 6).


The humerus and femur are only slightly longer in Hemiauchenia macrocephala compared to Palaeolama mirifica. The difference in length increases in the radius and tibia, and even more in the metacarpal, metatarsal, and proximal phalanges (Fig. 9). The latter have strong W-shaped muscle scars on their proximal end.

The ecology and diet of Hemiauchenia macrocephala have been extensively investigated. While it was long assumed that Hemiauchenia macrocephala was an herbivore like modern camelids, what kinds of plants it was eating was of interest to paleontologists because it would help them infer the environment(s) inhabited by Hemiauchenia macrocephala. Originally, tooth crown height and morphology were the primary source of information used to study ancient diets of fossil herbivores. The dental morphology and degree of hypsodonty found in camelids such as genera Hemiauchenia and Camelops in North America during the Plio-Pliestocene was thought to reflect a predominantly grazing diet of animals living in relatively arid grasslands. However, over the past 25 years, researchers have used multiple techniques to try to infer the diets and paleoenvironments of North American camelids. These include morphologic analysis of the teeth, skull, and limb bones, microwear and mesowear of the molars, and stable isotopes of carbon and oxygen preserved in tooth enamel. New morphologic studies on Hemiauchenia determined that it was more of a browser than a grazer (Dompierre, 1995; Dompierre and Churcher, 1996). This was later confirmed by a number of studies using stable isotopes from tooth enamel (Feranec and MacFadden, 2000; Feranec, 2003; Kohn et al., 2005; Yann and DeSantis, 2014). These studies revealed that the diet of Hemiauchenia macrocephala was much more complex than previously thought. It was capable of adapting to local environmental conditions and surviving as either a browser, a browse-dominated mixed-feeder, or a graze-dominated mixed feeder. For example, in a study of a population from South Carolina, Kohn et al. (2005) found that Hemiauchenia macrocephala was predominantly browsing during the winter, and predominantly grazing in the summer. Feranec and MacFadden (2000) and Feranec (2003) studied specimens from numerous Florida sites of Blancan, Irvingtonian, and Rancholabrean age. They found a significant shift to greater consumption of more grasses and sedges in Irvingtonian and Rancholabrean populations compared to Blancan ones. Also working with specimens from Florida, Yann and DeSantis (2014) found that Hemiauchenia macrocephala altered its diet in response to changes in climate, unlike other herbivores which were either consistently browsers (such as deer and tapirs) or consistently grazers (horses, mammoths).
Isotopic analyses have also provided clues to possible predators of Hemiauchenia macrocephala. Feranec and DeSantis (2014) investigated the possible predator-prey relationship of various extinct mammals at different trophic levels from the Florida fossil localities Leisey Shell Pit 1A and Inglis 1A using carbon isotopes. The two extinct carnivorans (predators) investigated were Canis edwardii [a coyote-like canid, or “early wolf” (Feranec and DeSantis, 2014)] and Smilodon gracilis (a leopard-sized saber-toothed cat). Among the herbivores studied were the llamas Hemiauchenia macrocephala and Palaeolama mirifica along with other species from the orders Artiodactyla, Perissodactyla, and Proboscidea. While the specific species the carnivores were eating cannot be determined directly, the isotopic value of the herbivores’ diets can be found in the tissue of the carnivores’ teeth. That is to say, carbon isotope analyses can determine what kinds of plants herbivores were eating, and by proxy, the diets of prey consumed by carnivores. Feranec and DeSantis (2014) determined that at Leisey Shell Pit 1A Canis edwardii consumed a higher percentage of herbivores within the mixed feeders category than Smilodon gracilis, suggesting Canis edwardii was potentially a greater threat for Hemiauchenia macrocephala than Smilodon gracilis. Inglis 1A revealed a different story where both Canis edwardii and Smilodon gracilis had similar diets and both fed proportionally more on grazers than on mixed feeders and browsers.
Sources
- Authors: Richard C. Hulbert Jr. and Natali Valdes
- Original Completion Date: June 6, 2015
- Editors’ Names: Natali Valdes and Richard C. Hulbert Jr.
- Last Updated On: June 6, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida