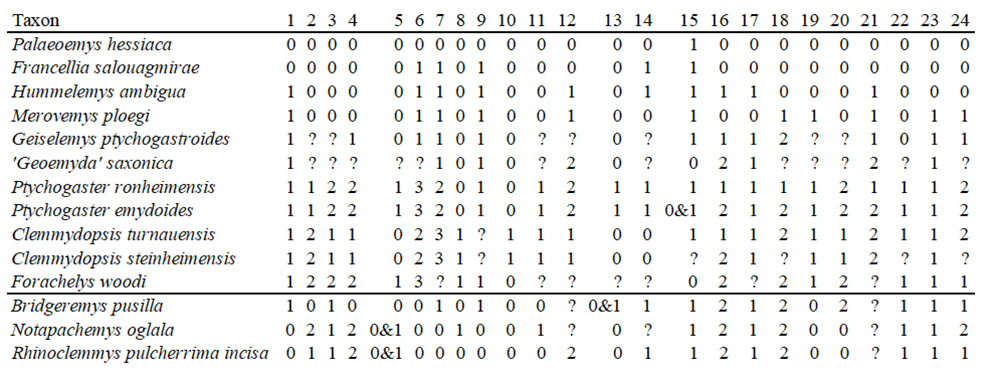
Abstract
Forachelys woodi gen. et sp. nov. is described from lower Miocene (Arikareean NALMA) exposures of the upper Las Cascadas Formation (~21 Ma) in the Panama Canal Basin. It represents the latest occurrence of ptychogastrine geoemydids in North America, a group known primarily from the early Eocene to late Miocene of Europe. Forachelys woodi shares features with taxa such as Geiselemys ptychogastroides (middle Eocene, Germany) and some species of the Ptychogaster/Temnoclemmys group (late Eocene to late Miocene, Europe). The epiplastral morphology is furthermore comparable to Bridgeremys pusilla (middle Eocene of Wyoming and Utah), a taxon previously interpreted as a rhinoclemmydine ancestor. Within the Las Cascadas Formation, F. woodi co-occurred with an undescribed species of rhinoclemmydine geoemydid that represents the oldest record of Rhinoclemmys (sensu lato). The Miocene fossil record of testudinoids in Central America is reviewed and reexamined. Testudinids are diverse in the Miocene of Panama and Honduras, represented by aff. Stylemys, Caudochelys (2–3 large to giant species), and an undescribed genus and species that resembles ‘Testudo’ costarricensis. Rhinoclemmydines persisted in Panama almost continuously from the early to late Miocene, with records from the Las Cascadas (~21 Ma), upper Culebra (~20–19 Ma), Cucaracha (~19 Ma), and lower Gatun (~12–11 Ma) formations. At least two concurrent rhinoclemmydines are present in the Cucaracha Formation of Panama, and two possibly concurrent taxa in the Gracias Formation (9–6.6 Ma) of Honduras.
Key words: Ptychogastrini, Rhinoclemmydinae, Geoemydidae, Testudinoidea, Miocene, Central America
https://doi.org/10.58782/flmnh.qvhz1455
Download Vol. 59, No.2, high resolution
Introduction
Today eight major groups of freshwater and terrestrial turtles are represented in Central America. These are the Chelydridae, Dermatemydidae, Kinosterninae, Staurotypinae, Rhinoclemmydinae, Testudinidae, Emydinae, and Deirochelyinae (Ernst and Barbour, 1989; Turtle Taxonomy Working Group, 2021). At least six of these groups have been reported in Central American Miocene fossil deposits, and two, the Podocnemididae and Trionychidae no longer live in the region (Laurito and Valerio, 2010; Cadena et al., 2012; Bourque, 2016). The testudinid ‘Testudo’ (or ‘Geochelone’) costarricensis Segura Paguaga, 1944, was described from the late Oligocene to early Miocene of Costa Rica based on a relatively complete shell (Segura Paguaga, 1944; Auffenberg, 1971; Coto and Acuña, 1986). The bulk of Miocene diversity has been collected in the Panama Canal Basin, where chelonians from the lower Miocene Culebra and Cucaracha formations have been previously reported (Cadena et al., 2012). The Culebra Formation (~23–19 Ma [Kirby et al., 2008; Buchs et al., 2019]) includes podocnemidids and trionychids (Cadena et al., 2012), and a new testudinid and rhinoclemmydine discussed later in this article. The Centenario Fauna (19.05–18.78 Ma [MacFadden et al., 2014]) has podocnemidids (at least two taxa), trionychids, Rhinoclemmys panamaensis Cadena et al. 2012, Rhinoclemmydinae (semi-aquatic/semi-terrestrial species), Staurotypus moschus Cadena et al. 2012, and testudinids (Cadena et al., 2012; Bourque, 2016; pers. observ.). The Cucaracha fauna is reexamined later in this article. Cheloniid sea turtles are known from the upper Miocene Gatun Formation (~12–8 Ma [Collins et al., 1996]) of Panama (Cadena et al., 2012). The nearshore marine Alajuela Formation (~9.77 Ma) of Panama contains the podocnemidid Bairdemys n. sp., trionychids, testudinids, and cheloniids (Bourque, in prep.; Bourque, 2016; MacFadden et al., 2017). Rhinoclemmys, a testudinid (‘Geochelone’), and a tricarinate kinosternine similar to members of the Kinosternon scorpioides (Linnaeus, 1766) complex were reported from the early Hemphillian (9–6.6 Ma) Gracias Formation of Honduras (Webb and Perrigo, 1984; Bourque, 2012). The Gracias turtle fauna is reevaluated later in this article. A trionychid, Apalone sp., emydine and deirochelyine emydids, and xerobatine testudinids were reported from the Mio–Pliocene (Hemphillian) of Costa Rica (Laurito et al., 2005; Laurito and Valerio, 2010). Most of the above-mentioned taxonomic groups dispersed south from more northerly latitudes in North America, with the likely exception of podocnemidids that may have dispersed northward from South America (Cadena et al., 2012; Weems and Knight, 2013). This article describes a new taxon from the El Lirio Norte locality, Panama, and introduces new early to late Miocene chelonian records from the Panama Canal and Honduras.
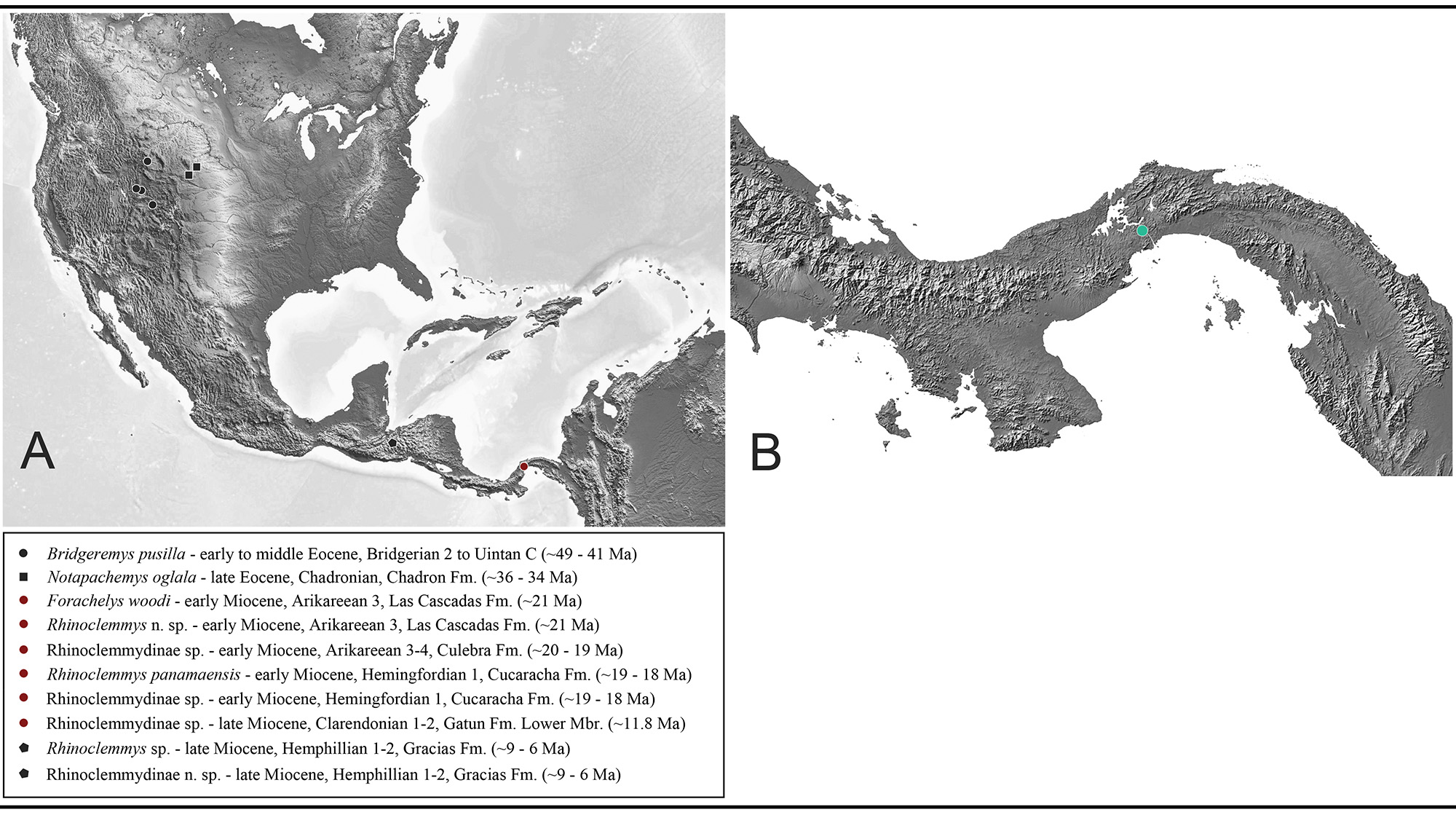
Locality Background
El Lirio Norte is located on the bank of the Panama Canal and preserves fossils from the lower Miocene (Arikareean NALMA) Las Cascadas Formation (~21 Ma) (Figs. 1–2), a volcaniclastic sequence that represents the oldest appearance of terrestrial habitats in the southern Panama Canal Basin (Rincon et al., 2012; Rincon et al., 2013; Bourque et al., 2013; Rincon et al., 2015; Buchs et al., 2019). Fossils from El Lirio Norte were collected between ca. 2009–2016 during expansion of the Panama Canal. The construction project exposed fresh rock facies of the upper Las Cascadas Formation. Field crews from the Florida Museum of Natural History (FLMNH) and the Smithsonian Tropical Research Institute (STRI) collected extensively at El Lirio Norte, which led to significant finds such as the first North American monkey Panamacebus transitus (Bloch et al., 2016), the floridatraguline camels Aguascalientia panamaensis and Aguascalientia minuta (Rincon et al., 2012), the protoceratid Paratoceras aff. P. tedfordi (Rincon et al., 2015), the anthracothere Arretotherium meridionale (Rincon et al., 2013), the flying squirrel Petauristodon sp. (MacFadden et al., 2014), a boid snake (‘cf. B. constrictor’) (sensu Head et al., 2012), and at least two crocodilians consisting of a cf. caimanine and longirostrine type (Hastings et al., 2013). Much has yet to be described from this diverse assemblage, collectively referred to as the Lirio Norte Local Fauna (LF) (Rincon et al., 2015). Chelonians from the Las Cascadas Formation at El Lirio Norte are described later in this article (see Discussion and Conclusions).

Materials And Methods
Virtual mesh models of UF 280569 and UF 280152 were produced using photogrammetry with a Canon EOS RP in the Division of Digital Imaging, Florida Museum of Natural History (FLMNH). Some figure images were developed from the 3D models where noted. Mesh models and image series are publicly available at https://www.morphosource.org (Table 1). UF 280569 (carapace and plastron with scute sulci outlined)—mesh: 000449207, image series: 000449204. UF 280569 (carapace and plastron)—mesh: 000449331, image series: 000449299. UF 280569 (plastron with partial carapace)—mesh: 000449334, image series: 000449302. UF 280152 (plastron)—mesh: 000449328, image series: 000449295. UF 280152 (right portion of anterior carapace)— mesh: 000449325, image series: 000449294.
Materials Examined
Las Cascadas Fm., Panama.—Forachelys woodi gen. et sp. nov.: UF 267054, UF 267127, UF 267136, UF 475212, UF 275421, UF 280152, UF 280422, UF 280569, UF 280570; Rhinoclemmys (El Lirio Norte): UF 280543, UF 532008; Podocnemididae (El Lirio Norte): UF 280542; Testudinidae aff. Stylemys (El Lirio Norte): UF 246844, UF 280339, UF 280568, UF 532151; Testudinidae gen. et sp. nov. (Empire West): UF 275495.
Cucaracha Fm., Panama.—Rhinoclemmys panamaensis: UF 237887, UF 257067; UF 280913, UF 281010; UF 281029, UF 281041; UF 281121; UF 281122; Rhinoclemmydinae sp. (semi-aquatic/semi-terrestrial): UF 281016; Rhinoclemmydinae indet.: UF 245513, UF 280539, UF 281031; Testudinidae: UF 257065.
Gatun Fm., Panama.—Rhinoclemmydinae sp.: UF 281207.
Gracias Fm., Honduras.—Rhinoclemmys sp. (thin-shelled): UF 17784, UF 18122, UF 18124, UF 46671, UF 532140, UF 532142, UF 532143, UF 532146, UF 532412; Rhinoclemmydinae n. sp. (thick-shelled): UF 17749, UF 17750, UF 18011; Rhinoclemmydinae indet.: UF 17743, UF 532144; cf. Emydidae: UF 532139, UF 532141, UF 532147; Caudochelys: UF 17785, UF 17786, UF 17791; Testudinidae (giant): UF 17790, UF 17792, UF 17793, UF 17795, UF 17796, UF 532145.
Other fossils.—Notapachemys oglala: UF 527000, UF 527218–UF 527223, UF 531033; Bridgeremys pusilla: UF 529329–UF 529331, UF 532057; UF 532058; Geiselemys ptychogastroides: GM 6970, GM XVIII 5000, GM XXXV 45 1962, GM XXXVI 30 62, GM XXXVI 204 62, GM XXXVI 238 62, XXXVIII 101 64, GM XXXVIII 148 67, GM XLIII 5 67; ‘Geoemyda’ saxonica: GM 6972; Hummelemys ambigua: GM 10537; Ptychogaster emydoides: FR 1081, FR 1083, FR 13896–FR 13970, FR 13992–FR 13994, FR 14019–FR 14038, FR 19479, FR 23030, FR 23032; SG 58, SG 66, SG 69.
Recent skeletons.—Rhinoclemmys annulata: UF/H 99676; Rhinoclemmys areolata: UF/H 54199, UF/H 76922; Rhinoclemmys funerea: UF/H 137638, UF/H 152560; Rhinoclemmys pulcherrima: UF/H 41046 (Nicaragua), UF/H 52797 (Costa Rica), UF/H 55442 (Honduras), UF/H 55574 (Honduras), UF/H 76923 (Guatemala); Rhinoclemmys rubida: UF/H 51411, UF/H 54197, UF/H 54787, UF/H 54980; Numerous Old World geoemydid skeletal specimens currently housed in the Division of Herpetology, FLMNH, were used extensively for comparison during this study.
Table 1. Mesh model hyperlinks for UF 280569 and UF 280152
Institutional Abbreviations
FR, Frick Collection, Department of Vertebrate Paleontology, American Museum of Natural History, New York; GM, Geiseltalmuseum Geologisch-Paläontologisches Institut der Martin-Luther Universität, Halle (Saale), Germany; SG, Département Histoire de la Terre, Muséum national d’Histoire naturelle, Paris; UF, Division of Vertebrate Paleontology, Florida Museum of Natural History, University of Florida, Gainesville, Florida; UF/H, Division of Herpetology, Florida Museum of Natural History, University of Florida, Gainesville, Florida.
Systematic Paleontology
Order TESTUDINES Batsch 1788
Family GEOEMYDIDAE Theobald 1868
Tribe PTYCHOGASTRINI De Stefano, 1903
FORACHELYS WOODI gen. et sp. nov.
Ptychogastrini gen. et sp. nov. (Las Cascadas Formation, Panama) Bourque, 2021
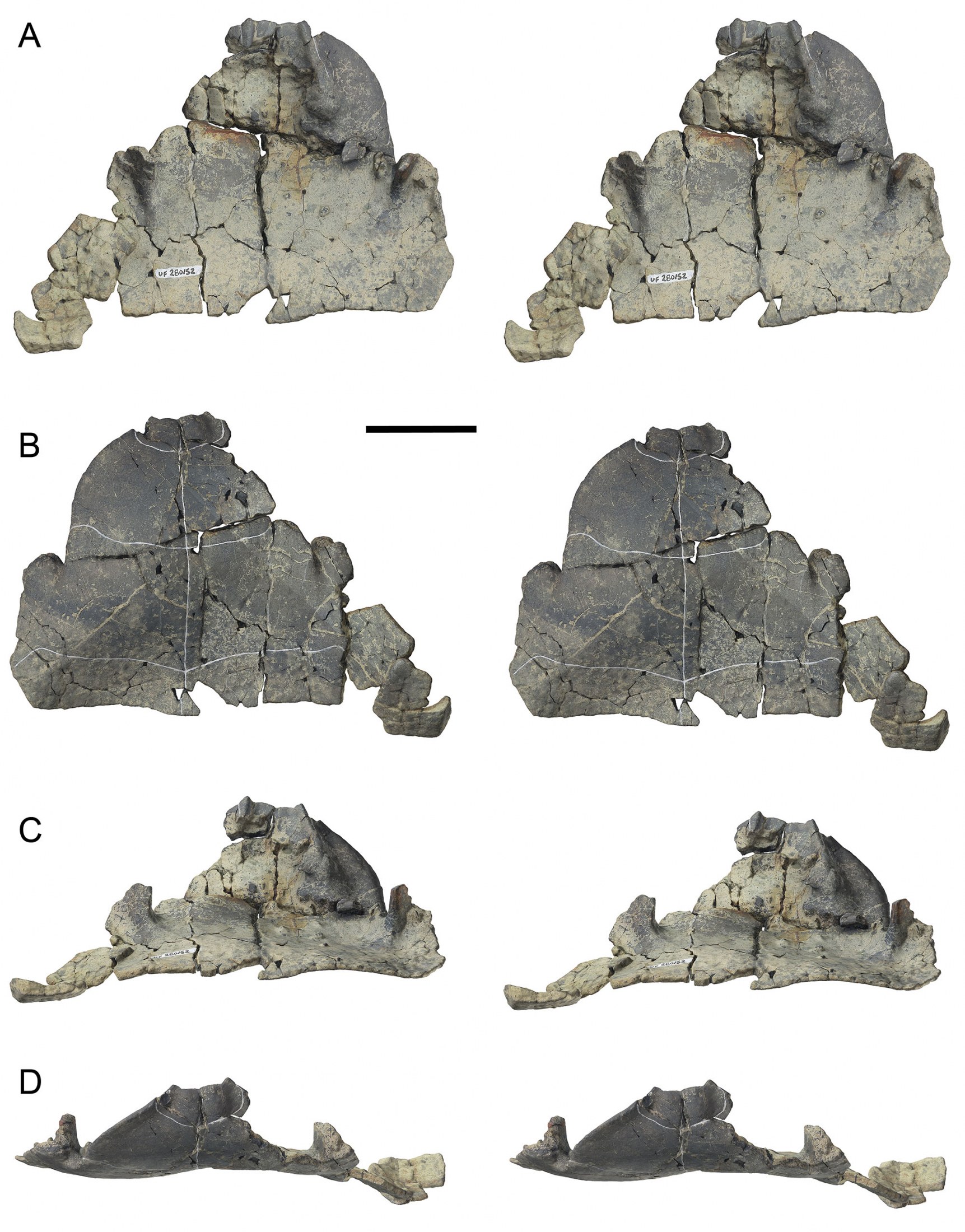
Holotype.—UF 280569, anterior carapace and plastron (Figs. 3–5A, 6–8). Collected by Aaron R. Wood, 14 November, 2011.
Paratype.—UF 280152, partial anterior carapace and plastral forelobe (Figs. 5B, 9–10). Collected by Aaron R. Wood, 4 May, 2012.
Referred Specimens.—UF 267054, entoplastron (Fig. 11G–H); UF 267127, right P7–8 fused (Fig. 11D–F); UF 267136, right P1; UF 275212, right P7; UF 275306, right P7 (subadult); UF 275421, right P6–7 fused; UF 280422, left P3; UF 280570, partial left hypoplastron (Fig. 11A–C); UF 532009, right P3–4 fused.
Zoobank Nomenclatural Act.—Genus: urn:lsid:zoobank.org:act:C59015F6-D3CF-483B-8EED-24FCEA2FDAB8. Species: urn:lsid:zoobank.org:act:1E23DB7D-92E7-42B6-B658- A4FC4B554751.
Etymology.—Generic epithet is combination of Latin foras for ‘outside’, and Greek chelys for ‘freshwater turtle’. Multiple derivations of foras are applicable to this taxon, such as Medieval Latin foresta for ‘forest’ referring to paleohabitat; Latin foris for ‘door’ or ‘gate’ referring to the movable plastral hindlobe; and Vulgar Latin foranus for ‘foreign’, ‘outsider’, or ‘outlander’ referring to the occurrence of a European turtle group at the southernmost tip of North America. Species epithet commemorates paleontologist Dr. Aaron R. Wood, friend and colleague who discovered and collected the type specimens.
Type Locality.—El Lirio Norte (YPA024), Panama Canal, Panama (Figs. 1–2).
Occurrence.—Early Miocene, Arikareean NALMA (Ar 3), Las Cascadas Formation, ~21 Ma (Tedford et al., 2004; Buchs et al., 2019).
Diagnosis.—Forachelys woodi is diagnosed by the unique combination of the following: anterior carapace (nuchal and N1) with tall rounded over crest; V1 long and narrow (much longer than wide); distinct cervical notch between M1 set along anterior carapacial rim; cervical scute and M1–M3 elongate and with extensive underlap (shared with Ptychogaster emydoides and Ptychogaster ronheimensis); gulars with extensive dorsal overlap and deep step at visceral junction (shared with Geiselemys ptychogastroides and Bridgeremys pusilla); posterodorsal margin of gulars notched medially and overhangs deep visceral concavity (shared with B. pusilla); presence of dorsally thick distal gular tubercles that do not extend significantly anteriorly (shared with some G. ptychogastroides); deep dorsal antero-posterior concavity along midline between gular tubercles (canal to accommodate head) (shared with some P. emydoides); gulars short ventrally and do not overlap entoplastron (shared with ‘Geoemyda’ saxonica and some P. emydoides); carapacial and plastral bones fused at sutures (in adults) with possible exception of hyo-hypoplastral suture and hypoplastron to carapace connections (shared with most P. emydoides, and Notapachemys oglala); axillary buttresses elongate and suturally united to carapace, inguinal buttresses short and ligamentously attached to carapace at P7–P8 jucture facilitating plastral hindlobe kinesis (shared with P. emydoides and P. ronheimensis).
Description And Comparisons
Carapace.—Only the anterior half of the shell is preserved in the holotype UF 280569 and paratype UF 280152 (Figs. 3–10). The posterior carapace is unknown. Forachelys woodi was small to mid-sized with an estimated carapace length of ~18–20 cm. Measurements for preserved portions of the types are presented in Table 2. The carapace is tall with a rounded over crest along the midline of at least the nuchal and N1. This anterior tent-like feature compares well with Rhinoclemmys rubida (e.g., UF/H 51411). On the ventral face of the nuchal region there are two thick swellings along the marginal-visceral transition that are much thicker than the comparatively thin costals and neurals (Fig. 8B–C). These swellings are similar to the buttressing seen in Bridgeremys pusilla, as well as Ptychogaster emydoides and Ptychogaster ronheimensis (Hervet, 2006). External surface bone is smooth on the holotype but the paratype has numerous growth annuli of low relief along the costals (Fig. 9B). The anterior peripherals are flat dorsally and not flared. The anterior carapacial rim lacks inter-marginal notches but there is a substantial cervical notch formed by the anteriorly pronounced M1 set (Figs. 3–5). The cervical scute is more elongate than is typical of other geoemydids and there is broad contact with V1. V1 is long and narrow and the lateral sulci are straight anteriorly and sinuous posteriorly (Figs. 3–5). V2 and V3 are partially preserved and appear to be less elongate than V1. The V1-M1 sulcus is distinctly posteriorly oriented, more so than in other ptychogastrines, species of Rhinoclemmys, B. pusilla, and Notapachemys oglala which all have a shorter M1 and broader V1 than F. woodi. The bridge peripherals have only a faint lateral keel (where the shell margin transitions along the bridge) (Fig. 7) similar to N. oglala and different from species of Rhinoclemmys which have a pronounced lateral keel.
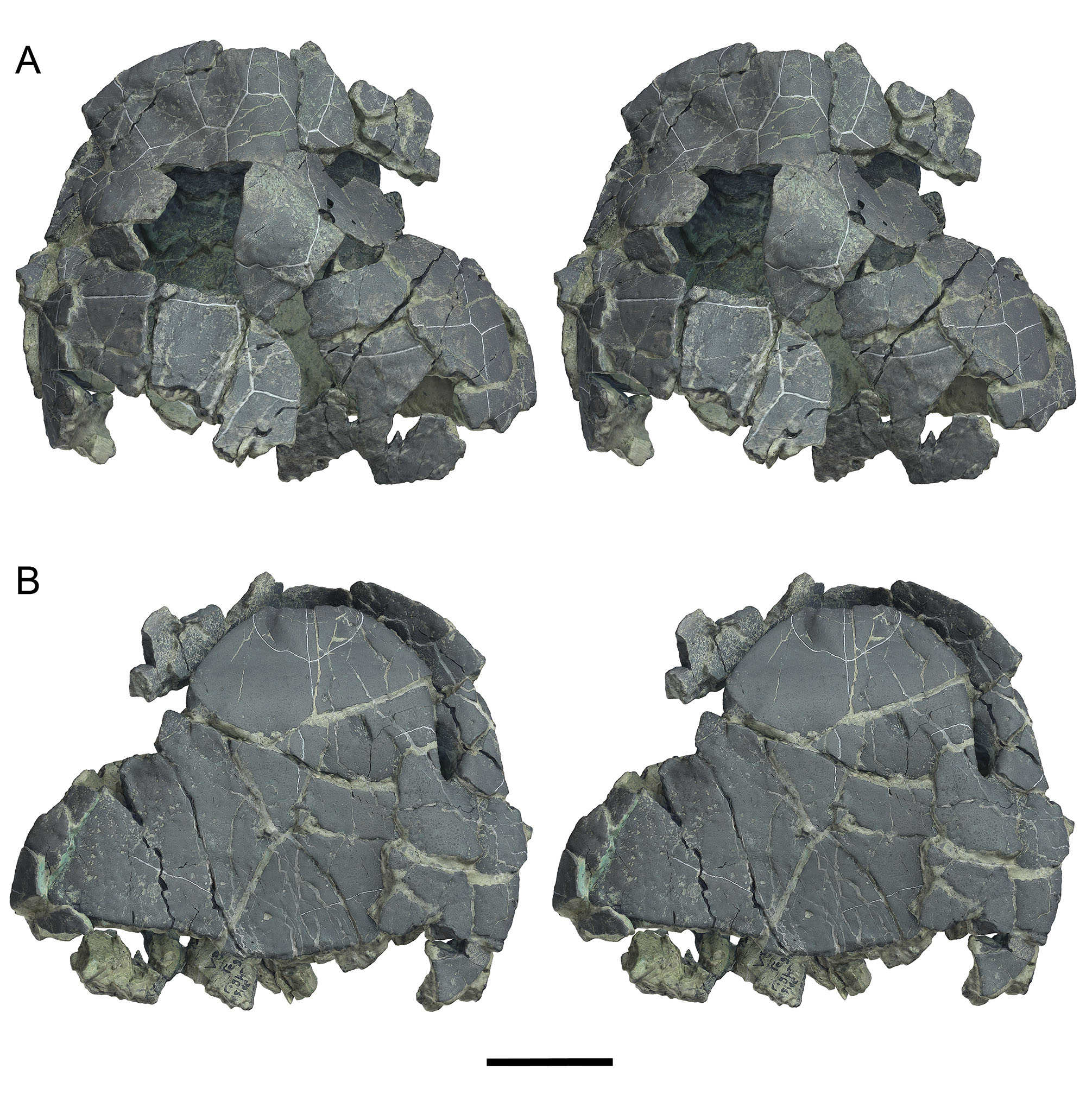
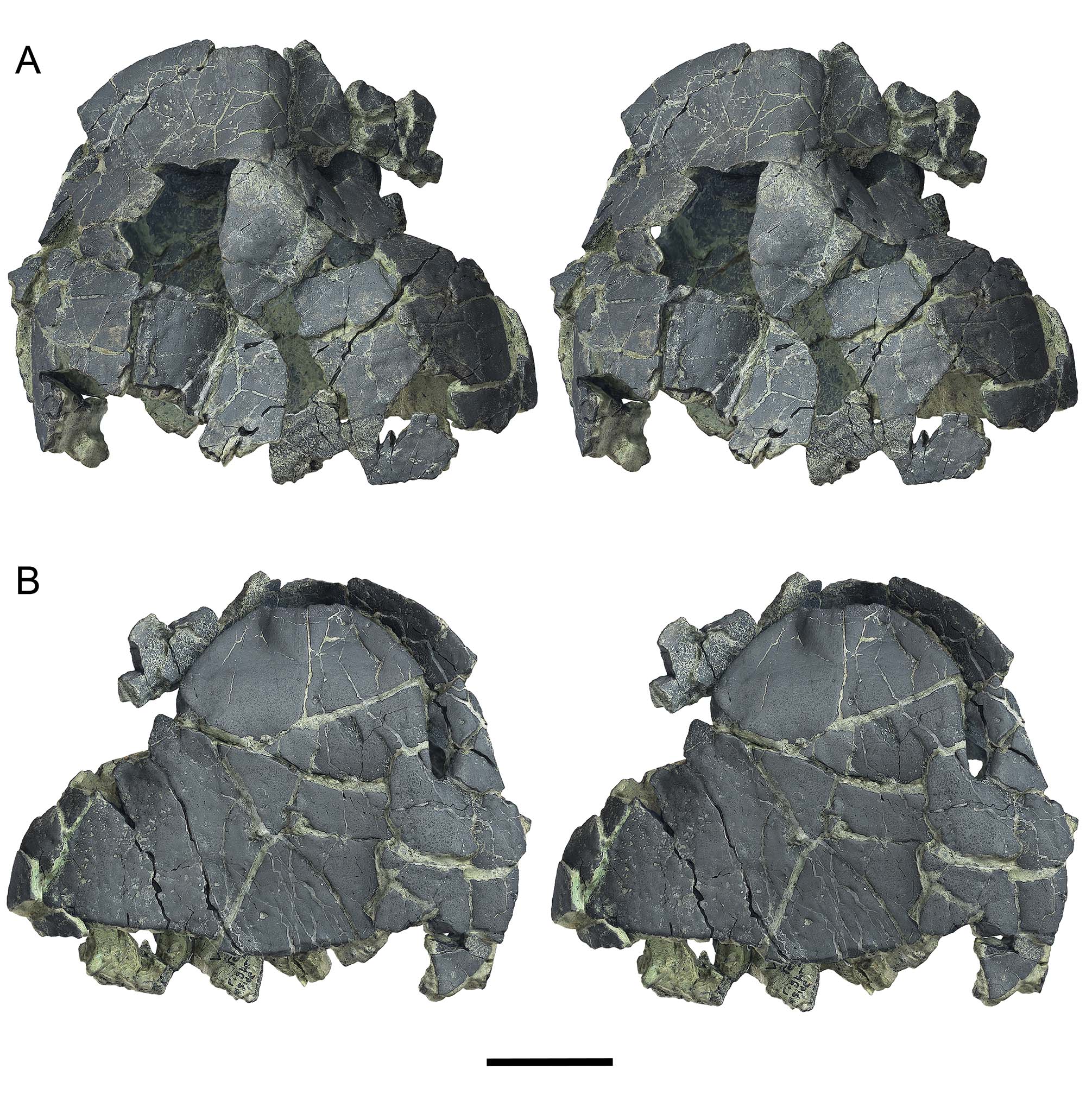
UF 280569 and UF 280152 are adults because all sutures are completely fused. Sutures are so ankylosed that peripheral, costal, neural, and nuchal contacts are indiscernible. This degree of ankylosis is similar to other box turtle morphotypes such as the ptychogastrines Ptychogaster emydoides Pomel (1847) and other members of the Ptychogaster/Temnoclemmys group (Bachmeyer and Schaffer, 1959; Schäfer, 2012) and extant emydine Terrapene. Most of the shell bones of Forachelys woodi are strikingly thin for a taxon that exhibits ankylosis, in particular the costals and hyoplastra. This differs significantly from Notapachemys oglala which also exhibits ankylosis but has much thicker shell bones overall.
The axillary bridge is akinetic and suturally united to the carapace. Its terminus is located on distal C1 (Figs. 8C–9A). The inguinal buttress is less extensive and its terminus appears to be located around the P7–P8 juncture, indicated on UF 267127, a fused right P7–P8 (Fig. 11D–F). The short inguinal sutural facet was clearly kinetic in life, having little or no interdigitating sutural teeth (Fig. 11D). Overall the shape and extent of the inguinal facet compares well to Ptychogaster emydoides. Axillary and inguinal musk ducts are not clearly discernible on the type and referred specimens, and this may be due to the fractured preservation of the fossils. However, in UF 280152 a deeply incised channel along the axillary buttress to P3 suture is possibly half of an axillary musk duct foramen (Fig. 9A), that in life would have been situated within the P3 hyoplastral suture.
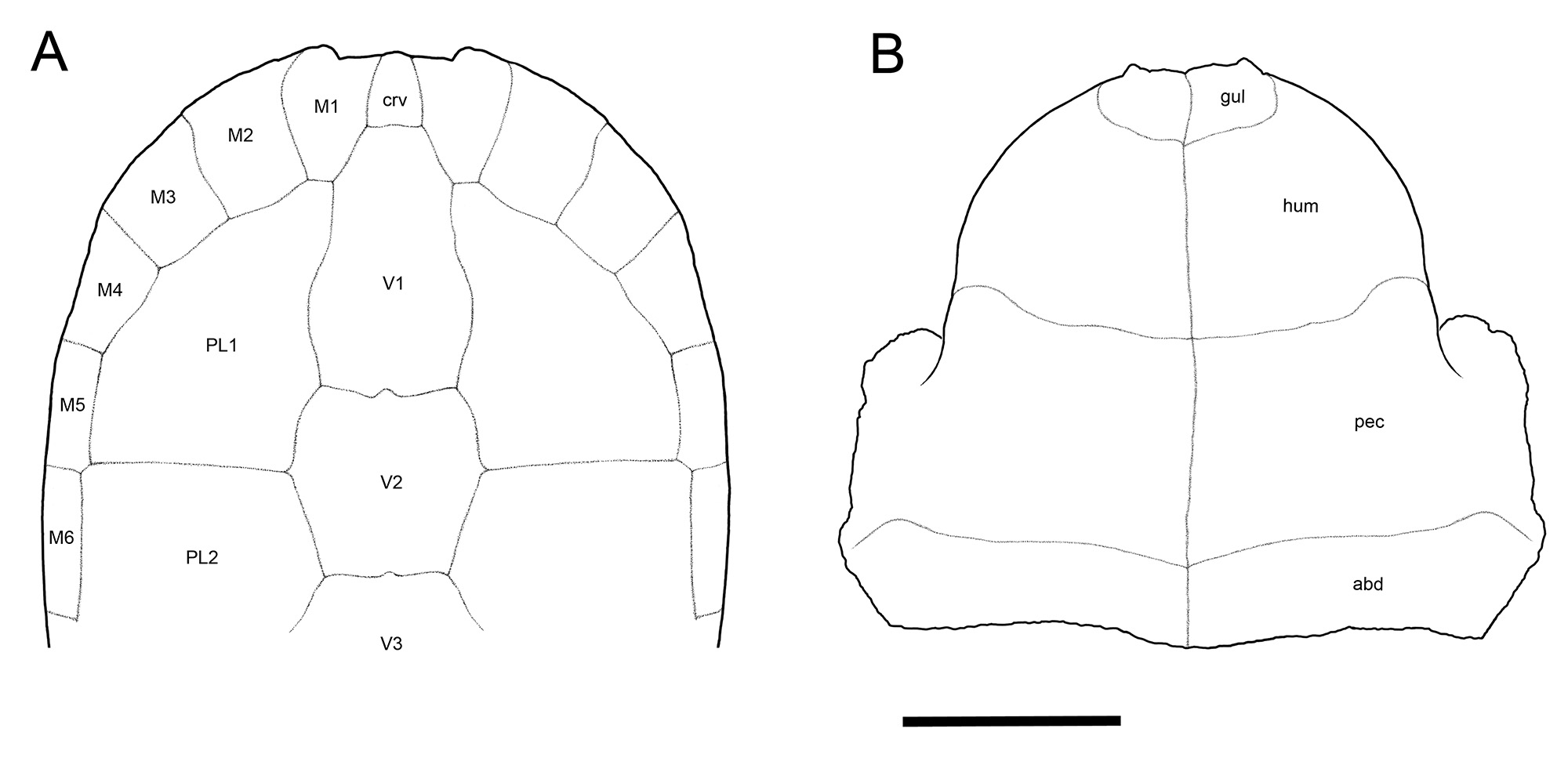
Plastron.—The holotype and paratype of Forachelys woodi preserve the plastral forelobe (Figs. 3–5, 10). Both are missing the paired hypo- and xiphiplastra, but a partial left hypoplastron (UF 280570) (Fig. 11A–C) is known. The plastron exhibits sutural fusion of the epi-, ento-, and hyoplastra, but the transverse hyo-hypoplastral suture was likely unfused. The unfused hyo-hypoplastral suture and kinetic junction between the carapace and plastral hindlobe may in part explain the missing preservation of the plastral hindlobe in both type specimens.
The epiplastra are the thickest portion of the shell (Figs. 8 and 10), distinct in morphology, and of all geoemydids examined most closely resemble Bridgeremys pusilla, and a number of ptychogastrines such as Geiselemys ptychogastroides, and species of the Ptychogaster/Temnoclemmys group such as P. laurae, P. buechelbergensis, P. boettgeri, and P. batalleri. There is a pronounced distal tubercle on the gular, which is present in most described species of ptychogastrines, i.e., ‘ptychogasterid spikes’ of Hervet (2004, 2006). The distal gular tubercle is notably thick dorsally, is wide posteriorly, and tapers to a fine point anteriorly, but projects only minimally from the anterior epiplastral margin (Fig. 8). There is a deep concave channel to accommodate the head and neck along the epiplastral midline formed between the inflated paired tubercles. This channel is particularly narrow in the paratype UF 280152, and overall narrower than observed in other ptychogastrines. The dorsal gular overlap is distinctly elongate. A deep step is present along the posterodorsal gular margin (Fig. 8) that is shared with B. pusilla, Notapachemys oglala, and many ptychogastrines including G. ptychogastroides, Ptychogaster emydoides. P. laurae, P. buechelbergensis, P. boettgeri, P. reinachi, P. ronheimensis, and P. batalleri. Likewise similar to B. pusilla, N. oglala, and the latter species of Ptychogaster, there is a medial notch along the posterodorsal gular margin along the visceral transition, and the posterodorsal gular margin overhangs a pair of deeply inset visceral concavites on the posteromedial epiplastra (Fig. 8B–C). In ventral aspect, the gulars are anteroposteriorly short and wide, and do not overlap the entoplastron. In fact, the posteroventral terminus of the gulars is well anterior to the anteromost entoplastral suture. This is difficult to observe due to ankylosis of the epi- and entoplastral sutures, however, can be seen most evidently in the paratype (UF 280152) (Fig. 10B). The humeral-pectoral sulcus overlaps transversely across the central portion of the entoplastron on UF 280152 where the sutural outline of the entoplastron is barely apparent due to some cracking along the fused sutural connection with the epi- and hyoplastra. An isolated entoplastron, UF 267054 (Fig. 11G–H), is attributed to F. woodi in that it is pyriform, elongate, relatively thin, lacks gular overlap, and the humeral-pectoral sulcus transversely crosses midway. An entoplastron with little to no gular overlap and a centrally positioned humeral-pectoral sulcus is seen in N. oglala, and a number of Ptychogaster species including P. (Temnoclemmys) batalleri, some P. buechelbergense, and P. emydoides. The humeral-pectoral and pectoral-abdominal sulci are anterodistally prominent, similar to other ptychogastrines. The pectoral-abdominal sulcus is situated well anterior to the hyo-hypoplastral suture (Figs. 3–5, 10) as in other ptychogastrines such as species of Ptychogaster and G. ptychogastroides. Conversely, the pectoral-abdominal sulcus intersects or almost intersects the hyo-hypoplastral suture medially in N. oglala, species of Rhinoclemmys, and B. pusilla. Just proximal to the margin of the forelobe is a ventral concavity at the distal gular-humeral junction that gives the forelobe margin a flared appearance (Fig. 6B–D). The entirety of the plastral bridge along the carapacial contact is dorso-ventrally flat and not steeply inclined, most evident in the paratype UF 280569 (Fig. 10).


The axillary buttresses are thin, moderately elongate, and suturally connected to the carapace (Figs. 8 and 10). The inguinal buttresses are significantly shorter than the axillary buttresses and attached to the carapace via a smooth kinetic facet evidenced by the fused P7–8 (UF 267127) (Fig. 11D–F) and isolated left hypoplastron (UF 280570) (Fig. 11A–C). The inguinal buttresses of Forachelys woodi are much shorter than those of Notapachemys oglala and species of Rhinoclemmys. Inguinal buttresses of some ptychogastrines such as Ptychogaster emydoides are similarly reduced to almost absent (Hervet, 2006). Also similar to P. emydoides, the left hypoplastron UF 280570 lacks a deep inguinal groove that is present at the base of the inguinal buttress on the ventral face of the hypoplastron in N. oglala and Bridgeremys pusilla. Semi-aquatic rhinoclemmydines such as Rhinoclemmys pulcherrima and Rhinoclemmys areolata have a similar combination of buttress to carapace articulations (axillary suturally anchored and inguinal kinetic) to that of F. woodi, species of Ptychogaster, and N. oglala, while more aquatic species like Rhinoclemmys panamaensis, Rhinoclemmys funerea, and Rhinoclemmys punctularia have rigid suturally anchored axillary and inguinal buttresses.


Discussion And Conclusions
Turtles from the Las Cascadas Formation
Chelonians from the Las Cascadas Formation at El Lirio Norte are previously unreported and include: Podocnemididae; Forachelys woodi (Geoemydidae, Ptychogastrini); Rhinoclemmys (sensu lato) (Geoemydidae, Rhinoclemmydinae); and two or three mid-sized testudinids (Figs. 12–13). The pleurodire is represented by a partial left portion of a carapace consisting of P8–9 and distal C5–7 (UF 280542) (Fig. 13K). UF 280542 compares well in size and morphology to extant Podocnemis examined (e.g., Podocnemis expansa UF/H 57965). The rhinoclemmydine is known from a partial disarticulated shell (UF 532008) (Fig. 12A–D) and an N1 or N3 (UF 280543) that represents a previously undescribed taxon. It differs from F. woodi in having a thicker unfused shell, wide V1, extensive akinetic inguinal buttresses, and epiplastra that lack significant dorsal gular overlap. The epiplastra instead strongly resemble species of Rhinoclemmys in having reduced distal gular tubercles, a low dorsal gular to visceral step, short dorsal gular overlap, ventral gular overlap on the entoplastron, a broad medial gular notch along the anterior gular margin, and a notched plastral forelobe margin at the gular-humeral sulcus (Fig. 11A–B). Rhinoclemmys n. sp. resembles more aquatic members of the genus such as Rhinoclemmys funerea (e.g., UF/H 152560) and Rhinoclemmys panamaensis (e.g., UF 237887) in having a thick relatively smooth acarinate shell (Fig. 11C). N1 is ovoid (not six-sided) as in Echmatemys, Bridgeremys pusilla, the holotype of R. panamaensis, and some R. funerea. The nuchal lacks marginal swellings on the visceral surface (Fig. 11B). The axillary and inguinal buttresses are elongate and sutured to the carapace. The inguinal buttress extends high onto C5 (Fig. 11D), more so than in extant species of Rhinoclemmys.
Table 2. Forachelys woodi measurements (in mm) of UF 280569 (holotype) and UF 280152 (paratype). Marginal measurements of UF 280569 for left side and of UF 280152 for right. Abbreviations: CERVL, cervical scute length; CERVW, cervical scute width; DGL, dorsal gular overlap length; FLL, plastral forelobe length; FLW, plastral forelobe width; GW, gular width; MGL, medial gular scute length; MHL, medial humeral scute length; MPL, medial pectoral scute length; M_L, marginal length; M_W, marginal width; V1L, Vertebral 1 scute length; V1W, Vertebral 1 scute width.
| Feature | UF 280569 | UF 280152 |
| CERVL | 12.72 | — |
| CERVW (posterior) | 7.89 | — |
| V1L | 43.99 | — |
| V1W (at M1) | 20.52 | — |
| M1L | 24.95 | — |
| M1W (anterior) | 16.88 | — |
| M2L | 23.91 | 28.99 |
| M2W (anterior) | 22.01 | 21.66 |
| M3L | 22.54 | 27.75 |
| M3W (anterior) | 22.12 | 17.9 |
| FLL | 86.04 | 85.83 |
| FLW (at axillary notches) | 68.74 | 71.8 |
| DGL | 25.32 | — |
| MGL | 14.5 | 12.65 |
| GW (right) | 12.92 | 12.9 |
| GW (left) | 13.98 | 12.09 |
| MHL | — | 27.1 |
| MPL | — | 38.01 |
One taxon of testudinid from El Lirio Norte has epiplastra with a low but pronounced gular to visceral step on the dorsal surface (Fig. 13E–I). The epiplastra are relatively thin for a testudinid, slightly concave distally on the ventral face, and the gulars have two short anteriorly projecting processes in presumed males UF 280568 (Fig. 13E–G) and UF 280339; a presumed female UF 246844 (Fig. 13H–I) lacks anterior projections and has a more rounded anterior gular margin. In these attributes and overall size and proportions, the epiplastra strongly resemble some specimens of Stylemys from the White River Group of the north central United States (Bourque et al., 2013). Stylemys or aff. ‘Stylemys’ gisellae has been previously reported from the late early Oligocene (Arikareean 1) Iniyoo local fauna (LF) of southern Mexico (Jiménez-Hidalgo et al., 2015; Jiménez-Hidalgo et al., 2021; Carbot-Chanona et al., 2022). A second testudinid taxon is known from only a single badly preserved partial shell (UF 275495) with a plastral forelobe unlike any other testudinid examined (Fig. 13A–D) and likely represents a new genus and species. The gulars are not anteriorly prominent and the posterodorsal gular margin is flush with the visceral transition (Fig. 13A and C). The plastral forelobe is conspicuously dorso-ventrally flat due to lack of inflated gulars. The flatness and shape of the plastral forelobe in ventral aspect resembles that figured for ‘Testudo’ costarricensis (Segura Paguaga, 1944; Auffenberg, 1971; Coto and Acuña, 1986) but I have not examined that taxon in person, and the dorsal epiplastra are currently unknown to me. The humeral-pectoral sulcus is situated just posterior to the ento-hyoplastral suture and does not broadly overlap the entoplastron as figured for costarricensis (Auffenberg, 1971; Coto and Acuña, 1986); however, it appears to contact the ento-hyoplastral suture at the midline (Fig. 13B and D).

Most chelonian fossils collected at El Lirio Norte are terrestrial and semi-terrestrial testudinoids, and overall diversity is low compared to the younger Centenario Fauna from the Cucaracha Formation (19–18 Ma) (MacFadden et al., 2014) which has more diversity of aquatic, semi-aquatic, and terrestrial chelonian taxa (Cadena et al., 2012). Chelonian fossils are also less abundant in the Las Cascadas Formation than from the Cucaracha Formation. Most of the fossils from El Lirio Norte are fragmentary and unfortunately most encased in extremely hard blue-green tuff that is often fused to the fossil surfaces with poor separation. Due to the difficulty of preparing these fossils, some specimens in the FLMNH collection remain encased in rock and currently unstudied.

The presence of a ptychogastrine in Panama at ~21 Ma is surprising but conceivable and represents a notable geographic range extension for this primarily European turtle group. A peak dispersal of Eurasian biota into North America occurred during the Late Oligocene Warming (~26–24 Ma) (Jiang et al., 2019) and between 21–18 Ma numerous Asian carnivores dispersed into North America (Qui, 2003). The anthracothere Arretotherium meridionale from the Las Cascadas Formation possesses similar dental characteristics as Oligo-Miocene Eurasian bothriodontine anthracotheres (Rincon et al., 2013). One of the oldest North American records of the Old World flying squirrel Petauristodon is from the Las Cascadas Formation (MacFadden et al., 2014; p. 129). It is possible that ptychogastrine turtles arrived in North America sometime in the late Oligocene to earliest Miocene and continued to disperse southward into the New World tropics, arriving in Panama by the early Miocene. Alternative hypotheses might include that Forachelys woodi is descended from proposed ptychogastrine or ptychogastrine-like stem rhinoclemmydines such as Bridgeremys pusilla and Notapachemys oglala which were already present in North America by the middle and late Eocene respectively (Hutchison, 1996; Hutchison, 2006; Bourque, 2021), or that multiple dispersals of ptychogastrines from Eurasia into North America have occurred from the Eocene to early Miocene. Ptychogastrines are not documented in the Asian fossil record so it is unclear if Beringia, the North Atlantic Land Bridge, or both were utilized as dispersal corridors from Europe into North America. Numerous examples of floral and faunal evidence suggest a North Atlantic connection between North America and Europe at times from the late Paleocene to the Oligocene or early Miocene (McKenna, 1975; Hably et al., 2000; Kvaček et al., 2000; Smith et al., 2006; Beard, 2008; Denk et al., 2011; Denk et al., 2012; Ellis et al., 2014; Jiang et al., 2019).
At least three of the chelonians represented in the Lirio Norte LF have North American affinities, with the likely exception of the podocnemidid that resembles Podocnemis from South America, and the enigmatic tortoise (UF 275495) that looks dissimilar to other North American testudinids except for the taxon costarricensis. The more common testudinid resembles Stylemys from the White River Group (Chadronian through Whitneyan) of northcentral North America. Stylemys or aff. ‘Stylemys’ has been previously reported from the late Oligocene (Arikareean 1) of southern Mexico (Jiménez-Hidalgo et al., 2015; Jiménez-Hidalgo et al., 2021; Carbon-Chanona et al., 2022). The oldest occurrence of a rhinoclemmydine in southernmost North America is Rhinoclemmys (sensu lato) (UF 532008) from the Lirio Norte LF. The oldest purported rhinoclemmydine ancestors are Bridgeremys pusilla and Notapachemys oglala from the middle to late Eocene respectively of the western-midwestern United States (Hutchison, 2006; Adrian et al., 2021; Bourque, 2021). During the Oligocene, southern North America was a plausible paleotropical refugium for more northern-latitude Eocene associated turtle groups (e.g., Stylemys group and rhinoclemmydines) in the cooler climate of the post-Eocene world (Woodburn, 2004).

The terrestrial landscape in the late Oligocene to early Miocene (~25–19 Ma) in Panama has been proposed as tropical North American rainforest (Jaramillo, 2018) and the presence of the early North American monkey Panamacebus at El Lirio Norte (Bloch et al., 2016) indicates that paleotropical forest was present at or nearby the site at 21 Ma. El Lirio Norte represents a terrestrial freshwater fluvial paleodeposit (Rincon et al., 2012) and previously reported components of the herpetofauna (Hastings et al., 2013), as well as still unreported taxa (pers. observ.) and aquatic to semi-aquatic turtles presented here support that interpretation. El Lirio Norte likely represents a paleotropical forest with freshwater streams and/or rivers amongst a volcanic complex of stratovolcanoes and volcanic domes (Buchs et al., 2019). Forachelys woodi is proposed as a tropical forest dwelling turtle that was likely semi-aquatic/semi-terrestrial and used relatively shallow freshwater bodies. The thin fused shell bones and extensive marginal and gular rims of the carapace and plastron are suggestive of this life history. Such extensive bony margins are typically amplified in life when overlain with even more anteriorly pronounced scutes. The anteriorly extensive marginal and gular rims superficially resemble the small tropical forest dwelling tortoises Kinixys erosa and Kinixys homeana (Ernst and Barbour, 1989), which have relatively slender heads and forelimbs that can be withdrawn deeply into the shell and protected by the pronounced margins of the anterior shell.
Cladistic Results
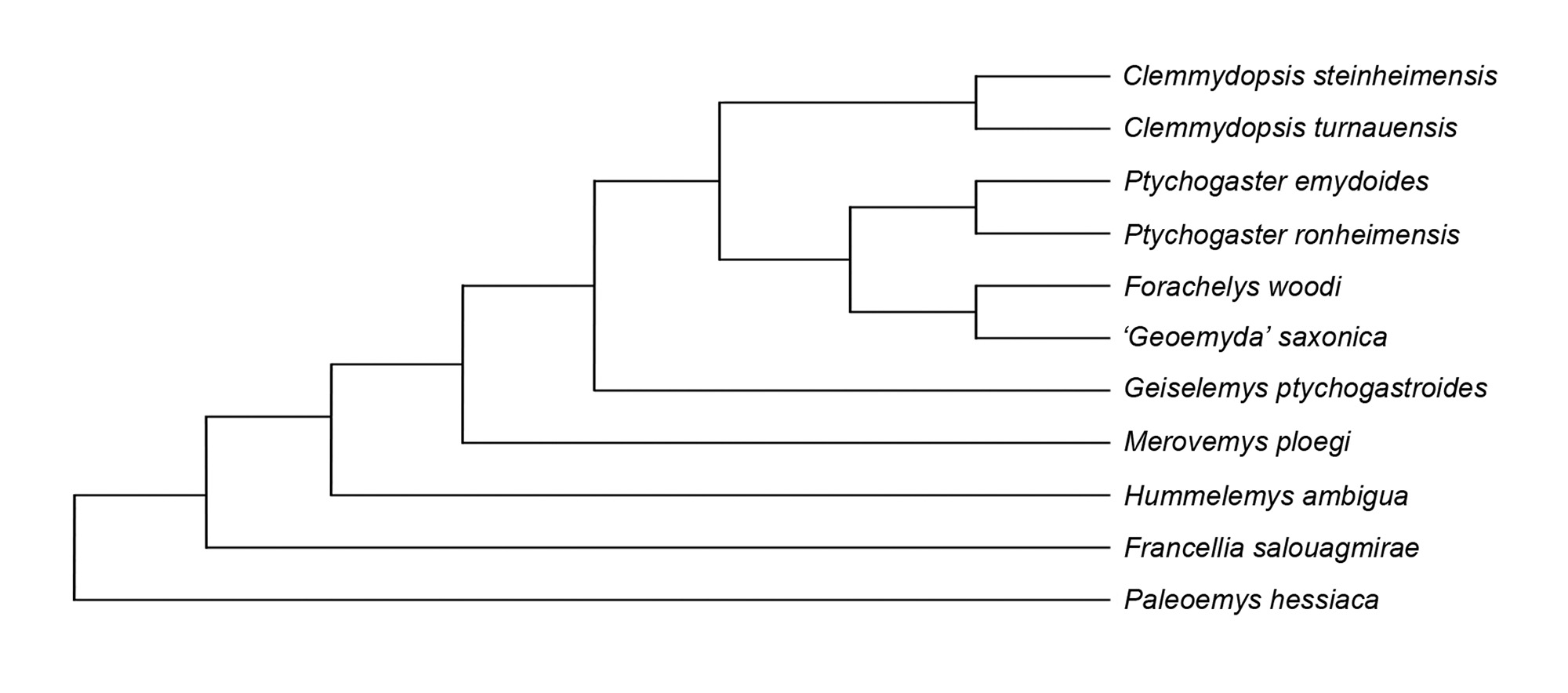
A parsimony analysis was conducted for proposed members of the Ptychogastrini with the matrix of Hervet (2006) (Appendix 1). Forachelys woodi was recovered nested within the Ptychogastrini and sister to the taxon saxonica (Fig. 14), which may be a junior synonym of Geiselemys ptychogastroides (Zimmermann-Rollius, 1966; Mlynarski, 1977; Hervet, 2006). This result is not unexpected considering similarities that F. woodi shares with G. ptychogastroides (Hummel, 1935) and the Ptychogaster/Temnoclemmys group. The epiplastra of F. woodi are most similar to numerous species of Ptychogaster, G. ptychogastroides and Bridgeremys pusilla. Hutchison (2006, p.75) first noted this similarity between the latter two taxa. A second analysis was performed during this study that incorporated F. woodi, Notapachemys oglala, B. pusilla, and Rhinoclemmys pulcherrima incisa into the matrix of Hervet (2006) (see Appendix 1), of which the position of F. woodi was consistently nested within the Ptychogastrini. I have chosen to omit the results here in that they are too preliminary and problematic regarding these additional taxa from a temporal perspective and in the absence of other pertinent geoemydids. Future work might build upon the observations of previous authors such as Garbin et al. (2019) and Hervet (2006), and further develop a detailed phylogenetically informative morphological matrix for the Geoemydidae that incorporates ptychogastrines and proposed stem and crown rhinoclemmydines.
Miocene Testudinoid Fossils in Central America
In an effort to find more records of ptychogastrines and better understand rhinoclemmydine diversity in the Miocene of Central America, I reexamined chelonian collections housed in the Division of Vertebrate Paleontology Collection at the FLMNH from the Panama Canal Basin and Gracias Formation of Honduras. Findings during that search are presented here (Figs. 15–18). The rhinoclemmydine record is rich in the Miocene of Central America and most localities examined have at least two co-occurring geoemydids or rhinoclemmydine morphotypes. Two geoemydid taxa co-occur in the Lirio Norte LF (~21 Ma). These are the ptychogastrine Forachelys woodi and a new species of rhinoclemmydine, tentatively identified here as Rhinoclemmys (sensu lato) (Fig. 12A–D). It is plausible that Rhinoclemmys n. sp. is ancestral to Rhinoclemmys panamaensis (Fig. 12E and G) in that they occur consecutively in the same region and both taxa resemble R. funerea (Fig. 12H) and the R. punctularia group (punctularia being the type species of Rhinoclemmys). The record of F. woodi from the Las Cascadas Formation is the latest known occurrence of ptychogastrines in North America and the New World.
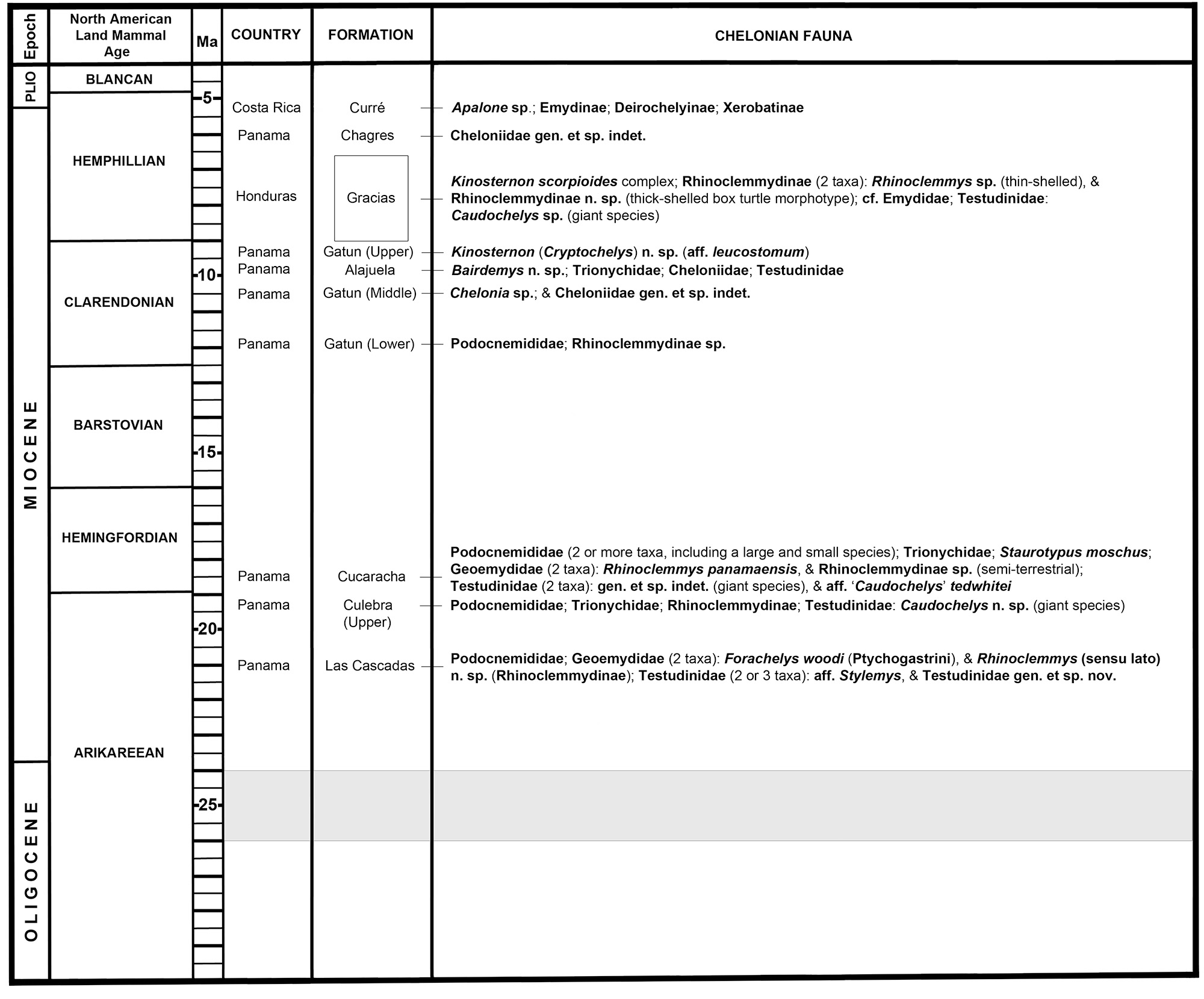
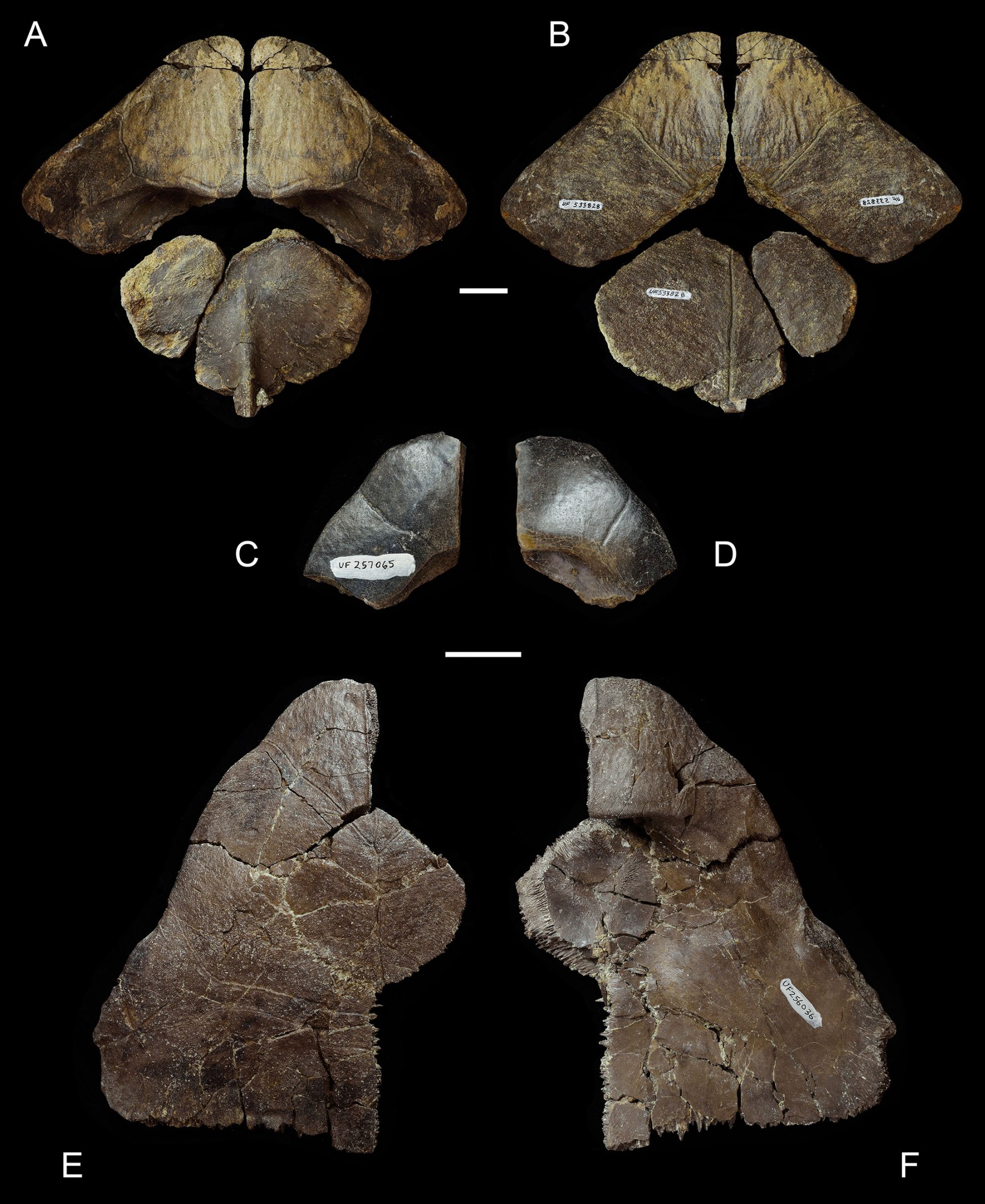
Turtles from the upper Culebra Formation (early Hemingfordian, ~20–19 Ma [MacFadden et al., 2014; Buchs et al., 2019]) include large podocnemidids and trionychids (Cadena et al., 2012). Previously unreported are large to giant sized testudinids (e.g., UF 242164, UF 257069, and UF 533828) (Fig. 16A–B) and a rhinoclemmydine (UF 242152) (Fig. 17C–D). The rhinoclemmydine is represented by a single partial left hypoplastron that is thick with an elongate, akinetic inguinal buttress.
The Centenario Fauna (19.05–18.78 Ma [MacFadden et al., 2014]) has two to three testudinoids (Cadena et al., 2012; pers. observ.) (Figs. 12E–G and 16C–D). At least two different rhinoclemmydines occur in the Centenario Fauna. Rhinoclemmys panamaensis (Fig. 12E and G) is a relatively large-bodied highly aquatic species similar in many aspects to Rhinoclemmys funerea (Cadena et al., 2012) (Fig. 12H), and the second taxon is a smaller semi-aquatic/semi-terrestrial rhinoclemmydine species with hindlobe kinesis (Fig. 12F). It is notable that two distinct rhinoclemmydine morphotypes with presumably diverse life history strategies co-occurred by this early appearance of Rhinoclemmys. Previously unreported from the Centenario Fauna is a mid-sized tortoise with epiplastra (e.g., UF 257065) (Fig. 16C–D) that look almost identical to ‘Caudochelys’ tedwhitei (Fig. 16E–F) from the approximately coeval Thomas Farm locality in north central Florida. The subgenus Caudochelys Auffenberg (1963) is used throughout this article at the generic level.

Fossil geoemydids are more fragmentary and lesser known in late Miocene deposits of Central America. In Panama, a single C1 (UF 281207) (Fig. 17A–B) that compares well to Rhinoclemmys is known from the Lower Member of the Gatun Formation (~11.8 Ma). On the visceral face of C1, the sutural contact for the axillary buttress is preserved. This suture is 23.31 mm long and is more proximally extensive than in most extant Rhinoclemmys examined, such as Rhinoclemmys pulcherrima and Rhinoclemmys funerea, where the axillary buttress terminates more distally on C1. However, the extensive reach of the axillary buttress in UF 281207 looks similar to Rhinoclemmys punctularia (e.g., UF/H 109148) from northeastern South America.
Turtle remains from the Gracias Formation of Honduras (~9–6.8 Ma) (Webb and Perrigo, 1984) are highly fragmentary (Fig. 18). The fauna is composed of multiple similarly aged localities (Webb and Perrigo, 1984). Two rhinoclemmydine taxa are present from the Gracias Formation, a thin-shelled taxon (cf. Rhinoclemmys) (Fig. 18A–D) and a thick-shelled box turtle morphotype (Fig. 18I–M). The latter undoubtedly represents a new taxon and resembles the emydine Terrapene in features of the nuchal and anterior peripherals. The thick-shelled taxon is known from the Rancho Lobo and El Carrizal localities (Webb and Perrigo, 1984) and possesses pronounced visceral swellings on the nuchal (Fig. 18J). The thin-shelled morphotype is present at Las Culebras, Tepusuna, and possibly Saddle localities (Webb and Perrigo, 1984) and lacks pronounced ventral swellings on the nuchal (Fig. 18B). Both forms have a medial keel or crest on the nuchal and broad V1 with PL1 sulci that barely overlap the nuchal (Fig. 18A and I). A posterior peripheral (UF 18124) (Fig. 18G–H) from Las Culebras may represent the thin-shelled taxon, but looks dissimilar to modern Rhinoclemmys examined. It is unclear which rhinoclemmydine taxon three peripherals (UF 17743; e.g., Fig. 18E–F) from the Saddle Locality represent but are relatively thin with distinct crenulated sculpturing along sulci and growth annuli. The P7 (Fig. 18E–F) has a tall broad inguinal sutural facet for hindlobe kinesis, upturned lateral marginal keel, and a musk duct foramen just posterior to the hypoplastral suture. Fragments of what appear to be a small emydid pond turtle were collected at Tepusuna and Las Culebras, but these are rare and poorly diagnostic past the tentative family level assignment here. Giant testudinids (Fig. 18N) occur at Tepusuna, and some specimens clearly resemble Caudochelys from more northern localities in North America and early Miocene of Panama, e.g., a subadult right P8 or P9 (UF 17791). However, other fragments are atypical for more northerly species of Caudochelys (e.g., Caudochelys crassiscutata from Florida) and could represent a second giant taxon or indicate variability of a single species. Two articulated posterior peripherals UF 17796 (Fig. 18N) are relatively thin and have broad scute sulci with raised sulcal borders, features that are atypical for Caudochelys but resemble some species of Chelonoidis or members of the Gopherus group.

Acknowledgments
This research was made possible by contributions from the following: Aaron Wood, Aldo Rincon, Bruce MacFadden, Jonathan Bloch, Carlos Jaramillo, Ed Stanley, Walter Joyce, Richard Hulbert, Coleman Sheehey, Tobias Landberg, Dick Franz, Dave Blackburn, Maria Vallejo, Lazaro Vinola Lopez, and Zach Randall. Smithsonian Tropical Research Institute (STRI), Authoridad del Canal de Panama (ACP) and Ministerio de Comercio e Industria (MICI) provided support and access to the Panama Canal Zone. Funding for this research was provided by the United States National Science Foundation (NSF) Partnerships in International Research and Education Grant (PIRE) 0966884 (OISE, EAR, DRL), EAR 0824299, and EAR 0418042, and the Department of Natural History, Florida Museum of Natural History (FLMNH). Photogrammetry models and uploads made in collaboration with Zach Randall (FLMNH). Walter Joyce, Evangelos Vlachos, Roger Portell, and Michal Kowalewski provided helpful manuscript comments. This is University of Florida Contribution to Paleobiology 883.
Literature Cited
Adrian, B., H. F. Smith, J. H. Hutchison, and K. E. B. Townsend. 2021. Geometric morphometrics and anatomical network analyses reveal ecospace partitioning among geoemydid turtles from the Uinta Formation, Utah. The Anatomical Record:1–35. doi.org/10.1002/ar.24792.
Auffenberg, W. 1963. Fossil testudine turtles of Florida. Genera Geochelone and Floridemys. Bulletin of the Florida State Museum 7(2):53–97.
Auffenberg, W. 1971. A new fossil tortoise, with remarks on the origin of South American Testudines. Copeia, 1971:106–117.
Bachmeyer, F., and H. Schaffer. 1959. Ein bemerkenswerter Schildkrötenfund (Ptychogaster grundensis nov. spec.) aus dem Untertorton von Grund, Niederösterreich. Annalen des Naturhistorischen Museums in Wien 63:78–89.
Batsch, A. J. G. C. 1788. Versuch einer Anleitung, zur Kenntniss und Geschichte der Thiere und Mineralien. Akademische Buchhandlung, Jena, 528 p.
Beard, K. C. 2008. The oldest North American primate and mammalian biogeography during the Paleocene–Eocene Thermal Maximum. PNAS 105(10):3815–3818.
Bloch, J. I., E. D. Woodruff, A. R. Wood, A. F. Rincon, A. R. Harrington, G. S. Morgan, D. A. Foster, C. Montes, C. A. Jaramillo, N. A. Jud, D. S. Jones, and B. J. MacFadden. 2016. First North American fossil monkey and early Miocene tropical biotic interchange. Nature 533:243–246.
Bourque, J. R. 2012. A fossil mud turtle (Testudines, Kinosternidae) from the early middle Miocene (early Barstovian) of New Mexico. Journal of Vertebrate Paleontology, 32:836–853.
Bourque, J. R. 2016. Side-necked turtles (Testudines, Pleurodira) from the ancient Gulf Coastal Plain of Florida during middle Cenozoic megathermals. Chelonian Conservation and Biology 15(1): 23–35.
Bourque, J. R. 2016 b. New mud turtles (Kinosternidae, Kinosternon) from the middle–late Miocene of the United States. Journal of Paleontology DOI: 10.1017/jpa.2015.63.
Bourque, J. R. 2021. A new geoemydid (Testudines, aff. Rhinoclemmydinae) from the upper Eocene Chadron Formation (White River Group) of northwestern Nebraska. Bulletin of the Florida Museum of Natural History 58(5):86–101.
Bourque, J., A. F. Rincon Burbano, A. Wood, J. Bloch, and B. MacFadden. 2013. New turtles (Reptilia, Testudines) from the Las Cascadas Formation, Panama Canal Basin, suggest low diversity in the early Miocene (Arikareean) neotropics. Journal of Vertebrate Paleontology (Supplement, Program and Abstracts) 33:91.
Buchs, D. M., D. Irving, H. Coombs, R. Miranda, J. Wang, M. Coronado, R. Arrocha, M. Lacerda, C. Goff, E. Almengor, E. Portugal, P. Franceschi, E. Chichaco, and S. D. Redwood. 2019. Volcanic contribution to emergence of Central Panama in the Early Miocene. Scientific Reports 9(1417):1–16.
Cadena, E. R., J. R. Bourque, A. F. Rincon, J. I. Bloch, C. M. Jaramillo, and B. J. MacFadden. 2012. New turtles (Chelonia) from the late Eocene through late Miocene of the Panama Canal Basin. Journal of Paleontology 86(3):539–557.
Carbot-Chanona, G., E. Jiménez-Hidalgo, J. A. Díaz-Cruz, G. Rivera-Velázquez, and V. H. Reynoso. 2022. A new large tortoise from the early Oligocene (Arikareean NALMA) of Oaxaca, southern Mexico and its phylogenetic position within Pan-Testudinidae. Historical Biology. doi.org/10.1080/08912963.2022.2114352.
Collins, L. S., A. G. Coates, W. A. Berggren, M-P. Aubry, J. Zhang. 1996. The late Miocene Panama isthmian strait. Geology, 24:687–690.
Coto, A. R. and R. A. M. Acuña. 1986. Filogeniade Geochelone costarricensis y la familia Testudinidae (Reptilia: Testudines) en el continente Americano. Revista de Biologia Tropical 34:199–208.
De Stefano, G. 1903. Ptychogaster miocenici della Francia conservati nel Museo di Storia naturale di Parigi. Palaeontographica Italica 9:61–94.
Denk, T., F. Grímsson, R. Zetter, and L. A. Símonarson. 2011. The biogeographic history of Iceland–The North Atlantic Land Bridge revisited. Pp. 647–668 in T. Denk, F. Grímsson, R. Zetter, and L. A. Símonarson, eds. Late Cainozoic Floras of Iceland. Topics in Geobiology, Vol. 35. Springer, Dordrecht.
Denk, T., F. Grímsson, and R. Zetter. 2012. Fagaceae from the early Oligocene of Central Europe: Persisting new world and emerging old world biogeographic links. Review of Palaeobotany and Palynology 169:7–20.
Ellis, D. and M. S. Stoker. 2014. The Faroe–Shetland Basin: a regional perspective from the Paleocene to present day and its relationship to the openeing of the North Atlantic Ocean. Pp. 11–31 in S. J. C. Cannon and D. Ellis, eds. Hydrocarbon Exploration to Exploitation West to Shetlands. Geological Society, London, Special Publication 397.
Ernst, C. H., and R. W. Barbour. 1989. Turtles of the World. Smithsonian Institution Press, Washington, D. C., 313 p.
Garbin, R. C., M. Böhme, and W. G. Joyce. 2019. A new testudinoid turtle from the middle to late Eocene of Vietnam. PeerJ 7:e6280 DOI 10.7717/peerj.6280.
Hably, L., Z. Kvaček, and S. R. Manchester. 2000. Shared taxa of land plants in the Oligocene of Europe and North America in context of Holarctic phytogeography. Acta Universitatis Carolinae–Geologica 44(1):59–74.
Hastings, A. K., J. I. Bloch, C. A. Jaramillo, A. F. Rincon, and B. J. MacFadden. 2013. Systematics and biogeography of crocodylians from the Miocene of Panama. Journal of Vertebrate Paleontology 33(2):239–263.
Head, J. J., A. F. Rincon, C. Suarez, C. Montes, and C. Jaramillo. 2012. Fossil evidence for earliest Neogene American faunal interchange: Boa (Serpentes, Boinae) from the early Miocene of Panama. Journal of Vertebrate Paleontology 32(6):1328–1334.
Hervet, S. 2004. A new genus of ‘Ptychogasteridae’ (Chelonii, Testudinoidea) from the Geiseltal (Lutetian of Germany). Comptes Rendus Palevol 3:125–132.
Hervet, S. 2006. The oldest European ptychogasterid turtle (Testudinoidea) from the lowermost Eocene Amber Locality of Le Quesno (France, Ypresian. MP7). Journal of Vertebrate Paleontology 26(4):839–848.
Hummel, K. 1935. Schildkröten aus der mitteleozänen Braunkohle des Geiseltales. Nova Acta Leopoldina 2:457–483.
Hutchison, J. H. 1996. Testudines. Pp. 337–338 in D. R. Prothero and R. J. Emry, eds. The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press, New York.
Hutchison, J. H. 2006. Bridgeremys (Geoemydidae, Testudines), a new genus from the middle Eocene of North America. Pp. 63–83 in I. G. Danilov and J. F. Parham, eds. Fossil Turtle Research Volume 1, Russian Journal of Herpetology 13 (Suppl.). St. Petersburg, Russia.
Jaramillo, C. 2018. Evolution of the Isthmus of Panama: Biological, paleoceanographic and paleoclimatological implications. Pp. 323–338 in C. Hoorn, A. Perrigo, and A. Antonelli, eds. Mountains, Climate and Biodiversity, First Edition. John Wiley and Sons, Ltd., New Jersey.
Jiang, D., S. Klaus, Y.-P. Zhang, D. M. Hillis, and J.-T. Li. 2019. Asymmetric biotic interchange across the Bering land bridge between Eurasia and North America. National Science Review 6:739–745.
Jiang, Y., M. Gao, Y. Meng, J. Wen, X.-J. Ge, and Z.-L. Nie. 2019. The importance of the North Atlantic land bridges and eastern Asia in the post-Boreotropical biogeography of the Northern Hemisphere as revealed from the poison ivy genus (Toxicodendron, Anacardiaceae). Molecular Phylogenetics and Evolution 139:106561.
Jiménez-Hidalgo, E., K. T. Smith, R. Guerrero-Arenas, and J. Alvarado-Ortega. 2015. The first Late Eocene continental faunal assemblage from tropical North America. Journal of South American Earth Sciences 57:39–48.
Jiménez-Hidalgo, E., E. B. Lander, I. Israde-Alcántara, N. W. Rodríguez-Caballero, R. Guerrero-Arenas. 2021. Earliest Arikareean (later early Oligocene) Iniyoo local fauna from Chilapa Formation of Santiago Yolomécatl area in northwestern Oaxaca, southern Mexico. Journal of South American Earth Sciences 109:1–18. 103307.
Kvaček, Z., S. R. Manchester, and H. E. Schorn. 2000. Cones, seeds, and foliage of Tetraclinis salicornioides (Cupressaceae) from the Oligocene and Miocene of Western North America: A geographic extension of the European Tertiary species. International Journal of Plant Sciences 161(2):331–344.
Kirby, M. X., D. S. Jones, and B. J. MacFadden. 2008. Lower Miocene stratigraphy along the Panama Canal and its bearing on the Central American peninsula. PLoS ONE 3(7):e2791. doi10.1371/journal.pone.0002791.
Laurito M., C., A. L. Valerio Z., L. D. Gómez P., J. I. Mead, E. A. Pérez G., and L. G. Pérez R. 2005. A Trionychidae (Reptilia: Testudines, Cryptodira) from the Pliocene of Costa Rica, southern Central America. Revisita Geológica de América Central 32:7–11.
Laurito M., C., and A. L. Valerio Z. 2010. Los Caballos Fósiles de la Formación Curré, Cantón de Coto Brus, Costa Rica. Museo Nacional de Costa Rica, San José, 131 pp.
Linnaeus, C. 1766. Systema Naturae, 12th ed. Laurentius Salvius, Holmia, 532 pp.
MacFadden, B. J., J. I. Bloch, H. Evans, D. A. Foster, G. S. Morgan, A. Rincon, and A. R. Wood. 2014. Temporal calibration and biochronology of the Centenario Fauna, early Miocene of Panama. The Journal of Geology 122:113–115.
MacFadden, B. J., D. S. Jones, N. A. Jud, J. W. Moreno-Bernal, G. S. Morgan, R. W. Portell, V. J. Perez, and A. R. Wood. 2017. Integrated chronology, flora and faunas, and paleoecology of the Alajuela Formation, late Miocene of Panama. PLoS ONE 12(1): e0170300. doi:10.1371/journal.pone.0170300.
McKenna, M. C. 1975. Fossil mammals and early Eocene North Atlantic land continuity. Annals of the Missouri Botanical Garden 62(2):335–353.
Mlynarski, M. 1977. Bemerkungen über die Schildkröten (Testudinoides) des Geiseltales. Eozäne Wirbeltiere des Geiseltales, Wissenschaftliche Beiträge Martin-Luther-Universität 2:99–105.
Pomel. A. 1847. Note sur las Mammifères et les Reptiles fossiles des terrains Éocènes de Paris, inferiéurs au dépôt gypseux. Archives des Sciences (Société de Physique et d’Histoire Naturelle de Genève) 4:326–330.
Qui, Z.-x. 2003. Dispersals of Neogene carnivorans between Asia and North America. Bulletin of the American Museum of Natural History 279:18–31.
Rincon, A. F., J. I. Bloch, C. Suarez, B. J. MacFadden, and C. A. Jaramillo. 2012. New floridatragulines (Mammalia, Camelidae) from the early Miocene Las Cascadas Formation, Panama. Journal of Vertebrate Paleontology 32:456–475.
Rincon, A. F., J. I. Bloch, B. J. MacFadden, and C. A. Jaramillo. 2013. First Central American record of Anthracotheriidae (Mammalia, Bothriodontinae) from the early Miocene of Panama. Journal of Vertebrate Paleontology 33(2):421–433.
Rincon, A. F., J. I. Bloch, B. J. MacFadden, and C. A. Jaramillo. 2015. New early Miocene protoceratids (Mammalia, Artiodactyla) from Panama. Journal of Vertebrate Paleontology 35(5):1–22.
Schäfer, D. 2012. Die schildkröten der gattung Ptychogaster Pomel, 1847 (Reptilia, Testudines). Dissertation. Ludwig Maximilian University of Munich, Germany. 387 p.
Segura Paguaga, A. 1944. Estudio de la primera especie nueve de tortuga fosil de Costa Rica con alguras generalidades sobre el orden testudines. La Esquela de Farmacia Costa Rica, 7 (73–74):9–29; (75–76):16–25; (77–78):13–24.
Smith, T., K. D. Rose, and P. D. Gingerich. 2006. Rapid Asia–Europe–North America geographic dispersal of earliest Eocene primate Teilhardina during the Paleocene–Eocene Thermal Maximum. PNAS 103(30):11223–112227.
Swofford, D. L. 2003. PAUP* Phylogenetic Analysis Using Parsimony (*and Other Methods) 4.0a (build 169).
Tedford, R. H., L. B. Albright III, A. D. Barnosky, I. Ferrusquia-Villafranca, R. M. Hunt Jr., J. E. Storer, C. C. Swisher III, M. R. Voorhies, S. D. Webb, and D. P. Whistler. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through early Pliocene epochs). Pp. 169–231 in M. O. Woodburne, ed. Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology. Columbia University Press, New York.
Theobald, W. 1868. Catalogue of the reptiles of British Burma, embracing the provinces of Pegu, Martaban and Tenasserim, with descriptions of new or little known species. Journal of the Linnean Society of London 10:4–68.
Turtle Taxonomy Working Group (Rhodin, A. G. J., J. B. Iverson, R. Bour, U. Fritz, A. Georges, H. B. Schaffer, and P. P. van Dijk). 2021. Turtles of the World: Annotated Checklist and Atlas of Taxonomy, Synonymy, Distribution, and Conservation Status (9th ed.). Chelonian Research Monographs, 8:1–472.
Webb, S. D., and S. C. Perrigo. 1984. Late Cenozoic vertebrates from Honduras and El Salvador. Journal of Vertebrate Paleontology, 4:237–254.
Weems, R. E., and J. L. Knight. 2013. A new species of Bairdemys (Pelomedusoides: Podocnemididae) from the Oligocene (early Chattian) Chandler Bridge Formation of South Carolina, USA, and its paleobiogeographic implications for the genus. P p. 289–303 in D. B. Brinkman, P. A. Holroyd, and J. D. Gardner, eds. Morphology and Evolution of Turtles. Vertebrate Paleobiology and Paleoanthropology Series, Springer, Dordrecht, Netherlands.
Zimmermann-Rollius, S. 1966. Beiträge zur Schildkrötenfauna der eozänen Braunkohle des Geiseltales. Hercynia N. F. 4:83–104.
Appendix 1