Abstract
We report abundant tortoise fossils from the Coralie Archaeological Site on Grand Turk Island (Turks Bank) and the non-cultural site of Indian Cave on Middle Caicos Island (Caicos Bank). These tortoises represent isolated populations of the extinct species Chelonoidis alburyorum, originally described from Abaco, The Bahamas. We describe the new material as two new subspecies, C. a. keegani on Grand Turk, and C. a. sementis on Middle Caicos. All fossil tortoises known thus far from The Bahamas and the Turks and Caicos Islands (= the Lucayan Archipelago) are likely referable to the polymorphic C. alburyorum. Lacking an epiplastral shelf distinguishes C. alburyorum from species of Chelonoidis in mainland South America and the Greater Antilles. Other diagnostic features of C. alburyorum are: extremely thin-shelled carapacial bones; flattened plastra (in presumed females only) or plastral concavities (mature males only), each with a series of horizontal boney ridges (bosses) on the internal surfaces; small to indistinct epiplas- tral apices; moderately sized to nearly absent xiphiplastral apices; and unique “bird-faced” entoplastral sculptures. A brief discussion follows on the possible sources, dispersal routes, environmental challenges, and ecological roles for the tortoises that colonized the West Indies.
Key words: Turks and Caicos Islands, Bahamas, Lucayan Archipelago, tortoise, Chelonoidis, fossil, zoo- archaeology, Holocene, new taxa.
Download Vol. 58, No.1, high resolution
Download Vol. 58, No. 1, low resolution
Introduction
The discovery of exquisitely preserved tortoise remains in Sawmill Sink, Great Abaco Island, The Bahamas, reawakened pursuit of the former diversity of West Indian tortoises, a quest that began a century and a half ago (Leidy, 1868), and continued with Williams (1950a, 1952), Auffenberg (1967, 1974), and Franz and Woods (1983). The remark- able Sawmill Sink sample, described as Chelonoidis alburyorum Franz and Franz (2009), consisted of complete shells and full sets of unbroken axial and appendicular bones. These fossils retained enough collagen for radiocarbon dating (Steadman et al., 2007, 2019; Steadman and Franklin, 2015) and even ancient DNA analyses, which established the genetic associations between C. alburyorum and extant species of Chelonoidis (Kehlmaier et al., 2017).
From the Dominican Republic, two other large, extinct tortoises recently have been described as new species: Chelonoidis marcanoi Turvey et al. (2017), based on fragmentary specimens from dry caves in Pedernales Province on the Barahona Peninsula; and C. dominicensis Albury et al. (2018), featuring an essentially complete individual (carapace, plastron, and nearly all axial and appendicular elements including a skull) from a flooded cave in La Altagracia Province, southeastern Dominican Republic. Chelonoidis dominicensis and other Greater Antillean fossil tortoises share more morphological traits with continental congeners than with the Lucayan C. alburyorum (Albury et al., 2018).
The fossils we now report from Grand Turk and Middle Caicos offer a new opportunity to appreciate tortoise diversity in the Lucayan region. They extend the known range of fossil Chelonoidis to some of the most remote islands of the expansive Lucayan Archipelago, where fossil tortoises are known now from 14 islands, although the morphology of most remains unstudied (Franz and Franz, 2009; Albury et. al., 2018; Steadman et al., 2019). Our goals here are: 1, to disclose a set of previously unrecognized or under-utilized characters on the internal surfaces of the shell; 2, to describe new subspecies of Chelonoidis alburyorum from the Turks and Caicos Islands; and 3, to discuss possible colonization routes for West Indian tortoises.
Localities
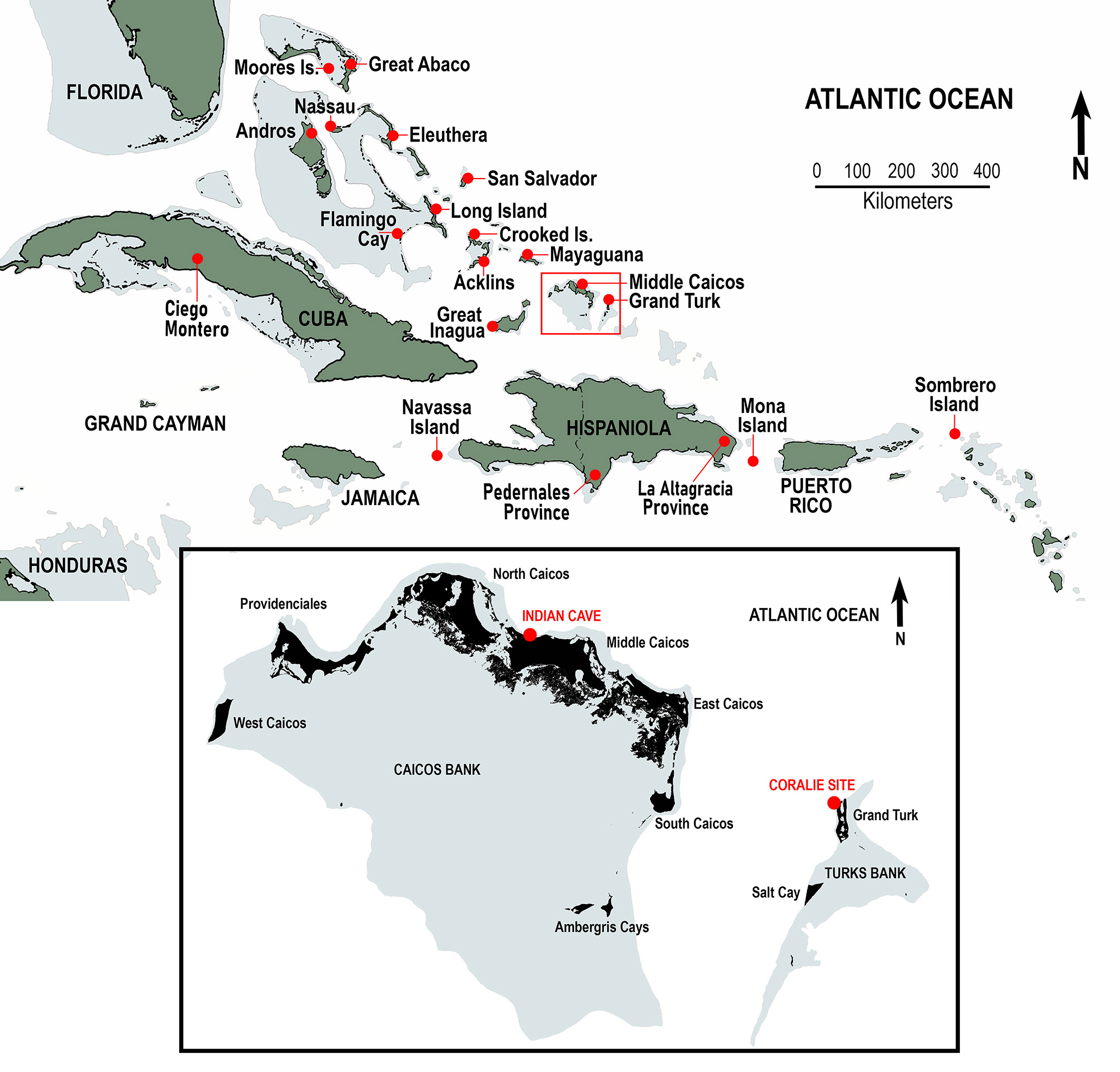
The Turks and Caicos Islands lie atop two separate carbonate banks, referred to as the Turks Bank and the Caicos Bank, both located southeast of the Great Bahama Bank, northeast of Great Inagua and Cuba, and north of Hispaniola (Fig. 1). These banks are separated by deep water of the Turks Island Passage, also known as the Columbus Passage. Grand Turk is the largest island on the Turks Bank (area 17.39 km2 or 6.71 mi2); Middle Caicos is the largest island on the Caicos Bank (area 144.2 km2 or 55.5 mi2). The distance between Grand Turk and Middle Caicos is 71.8 km (44.6 mi). The Turks and Caicos Islands (TCI) are politically independent from The Commonwealth of The Bahamas and are part of the British Overseas Territories. In this paper, we refer to The Bahamas and the TCI collectively as the Lucayan Archipelago or the Lucayan Islands, also known as the Bahamian Archipelago (e.g., Oswald and Steadman, 2018).
The Coralie Site (GT-3)
This account is taken primarily from Carlson (1999) and Carlson and Keegan (2004). The Coralie Archaeological Site lies along the western shore of the saline North Creek near the northern end of Grand Turk (Fig. 2A). A limestone ridge (elevations up to 26 m) parallels the length of the eastern (windward) shoreline, east of North Creek. Much of Grand Turk’s western side, including the archaeological site, is less than 9 m in elevation. GT-3 is located within 20 m of the creek’s mangrove fringe, only 1–2 m above sea level.
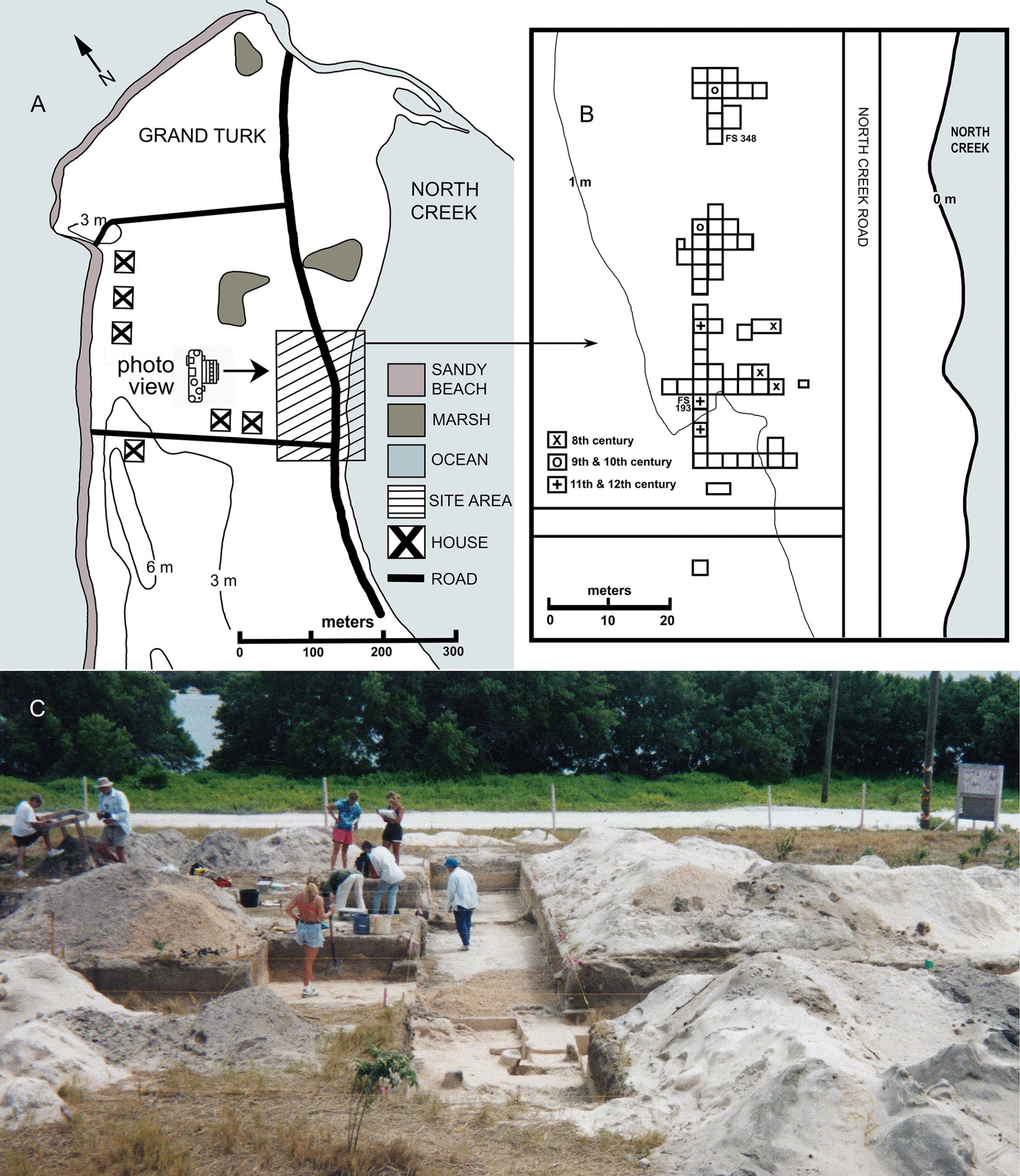
A land developer cleared the Coralie area to construct a housing subdivision in 1991, but the project never fully materialized. Brian Riggs, a TCI National Museum representative, was on-site during the land-clearing episodes to record the occurrences of cultural artifacts, including pottery and burnt conch shells. Based on Riggs’ report, William Keegan and Barbara Toomey visited the site in February 1992, digging 35 test pits with shovels.
From 1995 to 1997, field crews from the Florida Museum of Natural History, University of Florida, and the Turks and Caicos National Museum, systematically excavated the site under the direction of William F. Keegan and Lizabeth (Betsy) A. Carlson. Altogether, the archaeological teams at Coralie included over 100 volunteers, who contributed 10,000 person-hours (Carlson, 1999). Workers removed 271 m3 of sediment; excavations progressed in blocks of 2 x 2 m laid out in part as a long trench that paralleled North Creek (Fig. 2B–C). These excavations reached the water table at an average depth of 1 m. A series of perpendicular trenches were also dug that extended eastward from the primary trench. Sediment profiles showed dark, anthropogenic soils with many bone fragments, 10–40 cm thick, overlain by 25–65 cm of beach sand. The workers removed sediments with trowels and brushes in 10 cm vertical increments. Specimen recovery data were recorded as plot coordinates and then were assigned Field Specimen (FS) numbers.
GT-3 probably was not a full-time village but more likely a seasonal encampment for harvesting marine resources. The site’s food middens include abundant marine mollusks, marine fishes, and sea turtles in addition to tortoises, iguanas, and birds. The tortoise remains at GT-3 probably represent secondary victuals, taken sporadically during times of residency.
Ten conventional radiocarbon (14C) dates, mostly from charcoal samples from across the site, document two main periods of occupation at GT-3: an earlier one centered at cal AD ~700 to 900, and a later one at cal AD ~1000 to 1200 (Carlson, 1999; Steadman et al., 2019). The relatively early 14C dates are generally from the eastern side of the site, closest to North Creek. Two AMS 14C dates, each from a single bone of the extinct tortoise, are cal AD 771–903, 918–965 from FS 234, and cal AD 1042–1104, 1117–1222 from FS 82 (Steadman et al., 2019). These AMS 14C dates on tortoise bone correspond, respectively, with the older and younger time intervals originally reported by Carlson (1999).
To summarize, the Grand Turk tortoise was contemporary with the Amerindians (Ostionoid people) who initially occupied Coralie (site GT-3) at ~AD 700–800 (Keegan, 1997). These colonists ate tortoises, as evidenced by the burned tortoise bones excavated from cooking hearths. Tortoise bones also occur in the younger strata of GT-3 (cal AD ~1000–1200), where even more of the bones are charred.
Indian Cave (site MC-37)
Indian Cave is located adjacent to Dragon Cay Resort (formerly Blue Horizon Resort), west of Conch Bar, Middle Caicos Island. Its main entrance faces south in a prominent east-west trending limestone ridge, along the northern shore of the island, at an elevation of 9 m. East of Indian Cave, in the vicinity of Conch Bar and the airport, other dry caves occur in this ridge, which achieves elevations up to 42 m. The north face of the ridge is a series of headlands with cliffs that drop off to narrow beaches. Where undisturbed, the ridge sustains dry forest (coppice). Most of the southern half of Middle Caicos has lower relief and lies below 8 m elevation. The vegetation in these more poorly drained parts of the island consists of pineyard and mangroves.
With a walk-in entrance, Indian Cave features a single, well-lighted, cathedral-like room (Fig. 3). Multiple skylights penetrate the ceiling, providing sunlight and moisture to sustain vines and fig roots (Ficus) and rose apple (Clusia) trees that reach the cave sediments below. In 1998 and 2015, colonies of bats (Macrotus waterhousii) roosted in several small, short passages in the rear of the cave.
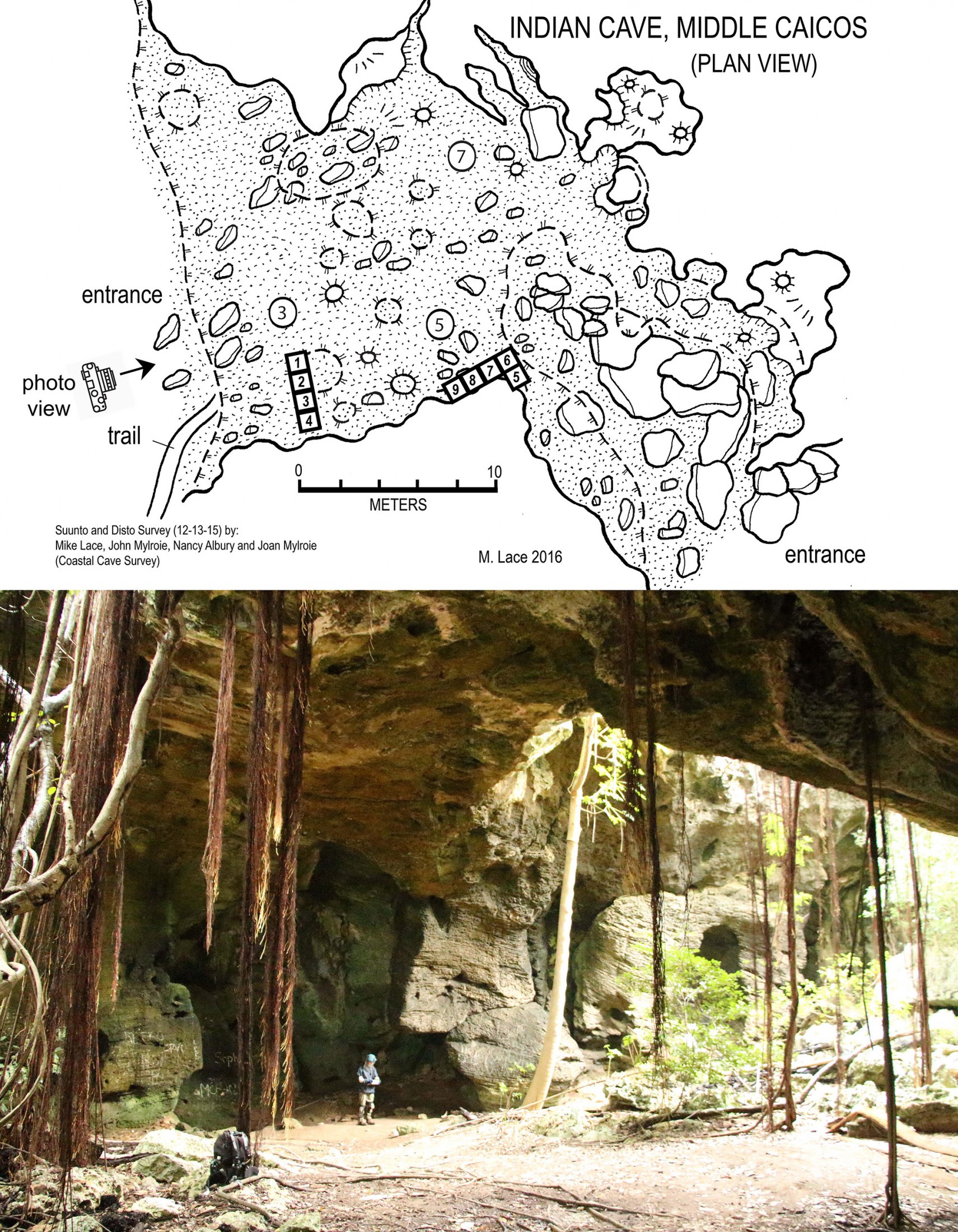
Indian Cave is the only prehistoric locality on the Caicos Bank with tortoise remains, which we recovered from non-cultural sediments. We excavated site MC-37 from 10–17 May 1998 (D. W. Steadman, W. F. Keegan, R. Franz, B. Riggs, A. V. Stokes, B. Toomey, and R. Toomey) and from 4–9 March 2001 (D. W. Steadman, J. Baker, S. Jones, P. O’Day, B. Riggs, and P. Sinelli). We discovered no evidence of prehistoric human occupation, although we obtained a very rich vertebrate record in a paleontological (non-cultural) context. Preliminary publications about Indian Cave are Franz (1999), Franz et al. (1999), Steadman (1999), and Carlson et al. (2006).
We excavated nine 1 x 1 m squares (referred to as Units 1–9) in two areas of Indian Cave (Fig. 3). Units 1–4 were connected to form a 1 x 4 m trench near the cave entrance inside the drip line, reaching a maximum depth of ~230 cm below surface or cmbs (which, on field labels, = ~300 cm below datum or cmbd); the other five units (5–9) were near the wall and much more shallow. Excavations followed natural stratigraphic breaks (Layers) that we removed in multiple Levels of 5–10 cm depth. The most productive unit for tortoises, Unit 7, next to the east wall of the cave, reached a depth of 90 cmbs. Unit 7 also yielded the most fresh-looking specimens. Site MC-37 yielded no complete shells of tortoises, nor much in the way smaller sections of shell that articulate with each other. The majority of tortoise bones from MC-37 were unassociated and incomplete. Most of the complete bones (limb elements, vertebrae) originated from on or near the sediment surface in Unit 7. They range from yellow to orange to brown, without charring, cut marks, or other evidence of tortoise-human interactions. Scattered small pieces of charcoal occurred in surface cave soils; a few bones from other Indian Cave units displayed minimal charring. The only human artifacts found in the cave were on the sediment surface, namely eight small pieces of Meillacan pottery, similar to those having African origins (possibly brought from Hispaniola), and a metal thimble similar to ones used for mending sails (Keegan, 1997).
For each catalogued specimen from excavation units 1–4 at MC-37, we provide provenience data, namely the excavation unit (Unit 1, 2, 3, etc.), the stratigraphic layer (Layer I, II, III, etc.), and the depth level (Level 1, 2, 3, etc.). Examples would be “(Unit 2, II/4)” or “(Unit 3, IV/17).” Excavations in units 5–9 proceeded much less meticulously; for these five units, we provide whatever data are available.
Tortoise fossils are the basis for the 14C chronology of Indian Cave (Steadman et al., 2019). The youngest AMS 14C date is cal AD 890–1020 from the surface of Unit 9. The other three finite AMS 14C dates on tortoise fossils from MC-37 range from cal AD ~650–770 to as old as cal AD ~10–130, although these dates are not in stratigraphic order. Five other tortoise bones from Indian Cave, ranging in depth from ~150 cmbd (~90 cmbs) to ~300 cmbd (~240 cmbs), did not have enough collagen to yield an AMS 14C date. The age of the deeper levels of our excavations at MC-37 probably is mid- to early Holocene. We found no stratigraphic breaks, or changes in fossil preservation, that were dramatic enough to suggest a transition with depth from Holocene to Pleistocene.
Methods and Materials
We directly compared the Turks and Caicos fossils with fossils of Chelonoidis alburyorum from Abaco and C. dominicensis from Hispaniola. We also compared the fossils with modern specimens of Geochelone sulcata, C. carbonarius, C. denticulatus, and C. chilensis, and multiple taxa of Chelonoidis from the Galápagos Islands, the latter variously evaluated as allopatric species, sub-species, or populations of C. elephantopus sensu lato or C. nigra sensu lato (Poulakakis et al., 2008, 2015; Olson and Humphrey, 2017). Comparative specimens are from the Florida Museum of Natural History, University of Florida (UF), the National Museum of The Bahamas (NMB), the Turks & Caicos National Museum (TCNM), the Smithsonian Institution (USNM), the St. Augustine Alligator Farm Zoological Park, and private collections.
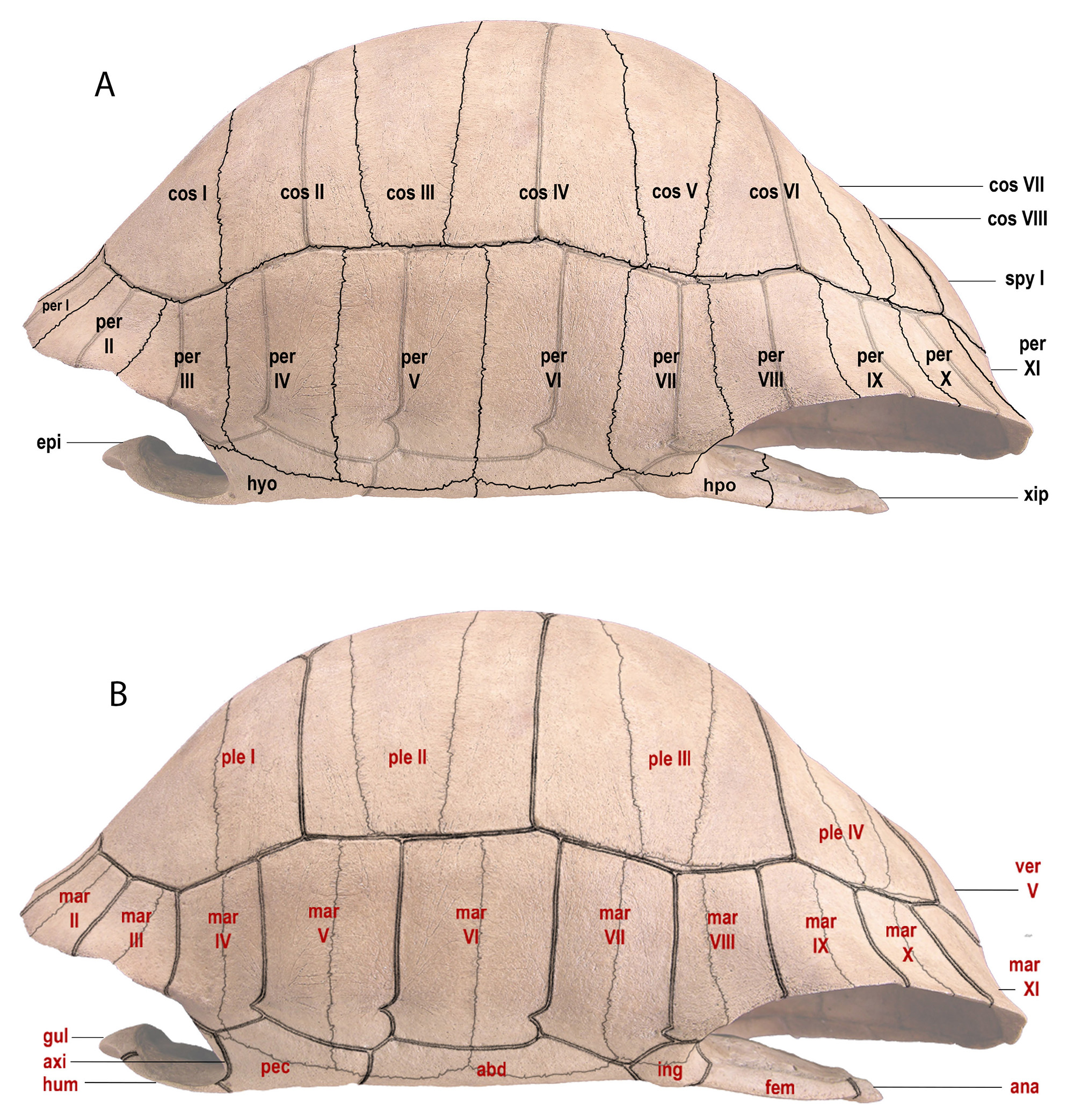
We generally follow the terminology used in Bramble (1971) and Franz and Franz (2009) for the names of carapacial and plastral bones, sutures, scutes, and sulci (Figs. 4–7). When possible, we took these measurements: CL, standard carapace (midline) length; PL, standard plastron midline length; GPL, greatest plastron length (including epiplastral and xiphiplastral apices); PW, width across the hyoplastral/hypoplastral suture. The terminology for limb bones follows Hulbert (2001). For excavation depths, cmbd = centimeters below datum; cmbs = centimeters below surface.
Internal Shell Characters
The skull, and external features of the carapace and plastron, traditionally have been used to define fossil tortoise taxa. Here we also evaluate internal sculptural features, such as muscle scarring on the internal surfaces of shells, as another source of characters to distinguish individual species of Chelonoidis. These skeletal features often register interactions between muscles, tendons, and skin, which together reflect functional and behavioral activities of the tortoise. Visual access to the internal surfaces of shells often requires extra preparation of fossils and modern specimens to expose these structures (Fig. 6). We suggest that evaluation of the following internal features is helpful to define taxonomic relationships in Chelonoidis.
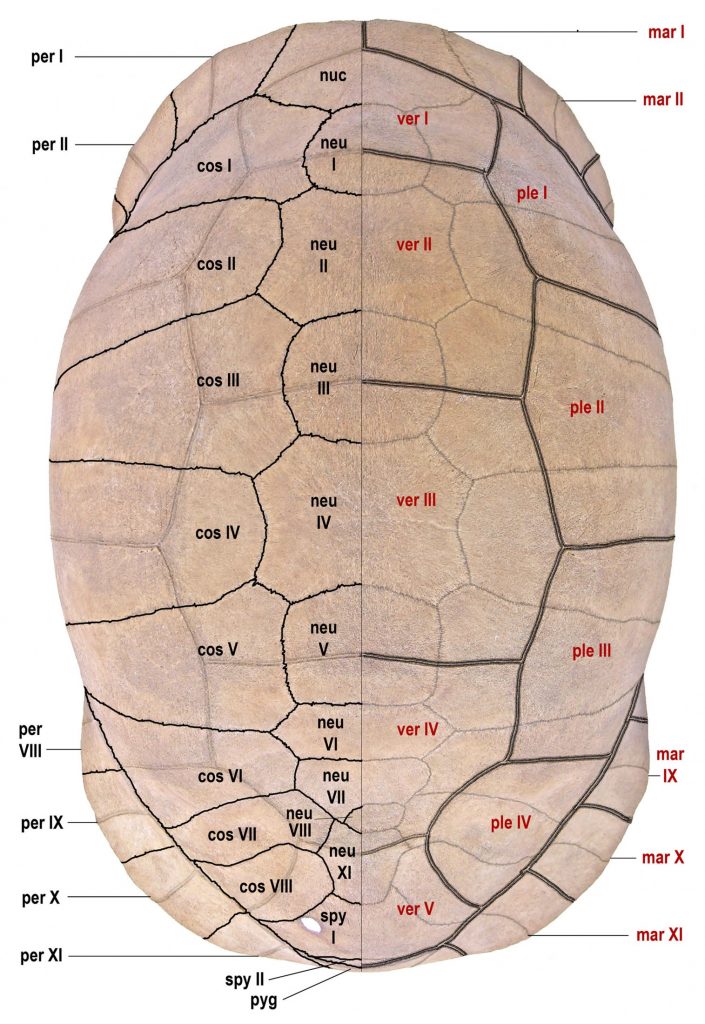
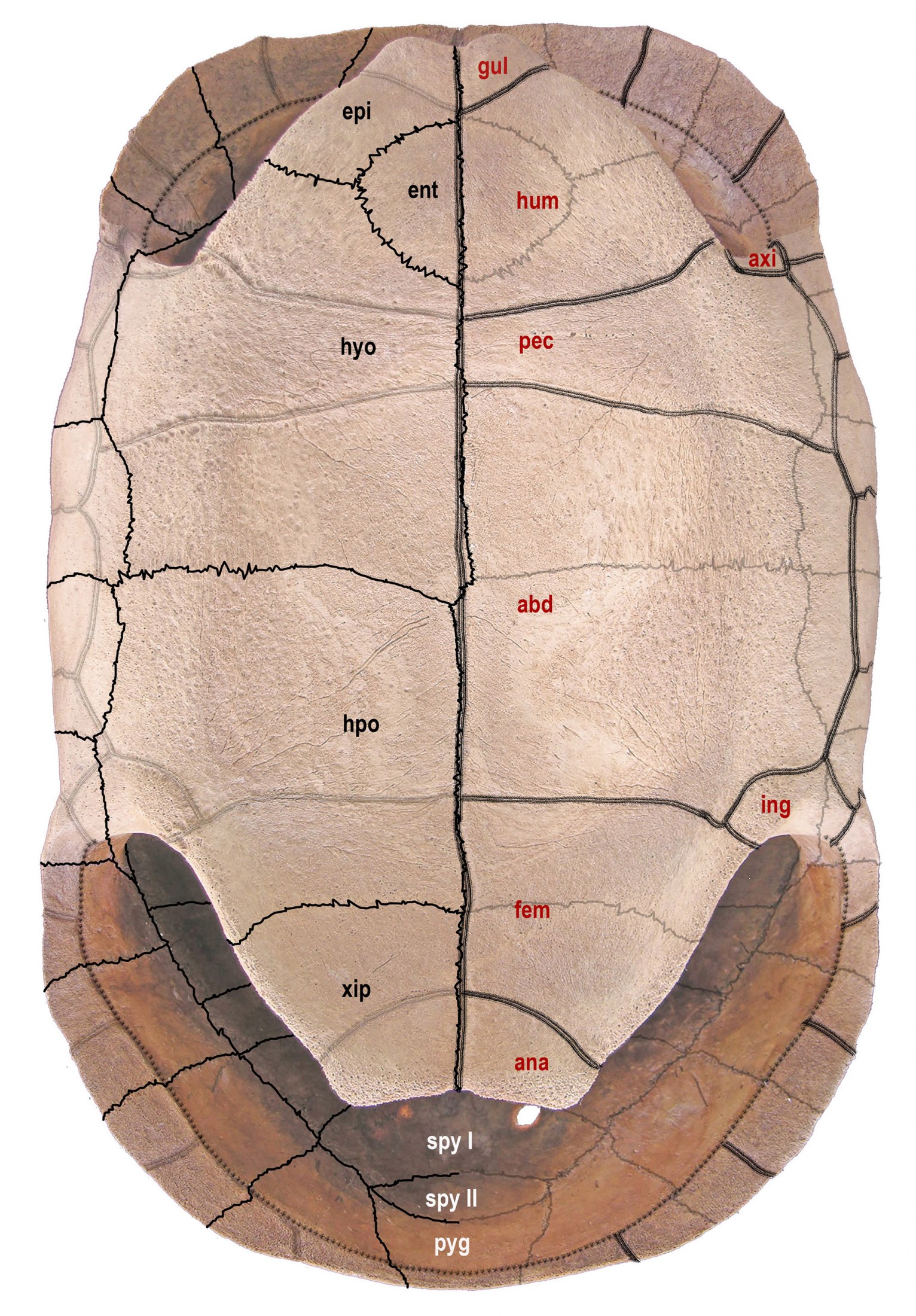
bones in black (on left) and scutes in red (on right). Single line = sutures; double line = sulci. Abbreviations
for bones and scutes apply to Figures 4–7. Carapace bones: cos, costal; neu, neural; nuc, nuchal plate;
per, peripheral; pyg, pygal; spy, suprapygal. Plastron bones: epi, epiplastron; ent, entoplastron; hyo,
hyoplastron; hpo, hypoplastron; xip, xiphiplastron. Carapace scutes: mar, marginal; ple, pleural; ver,
vertebral. Plastron scutes: abd, abdominal; ana, anal; axi, axillary; fem, femoral; gul, gular; hum,
humeral; ing, inguinal; pec, pectoral. Modified from Albury et al. (2018).
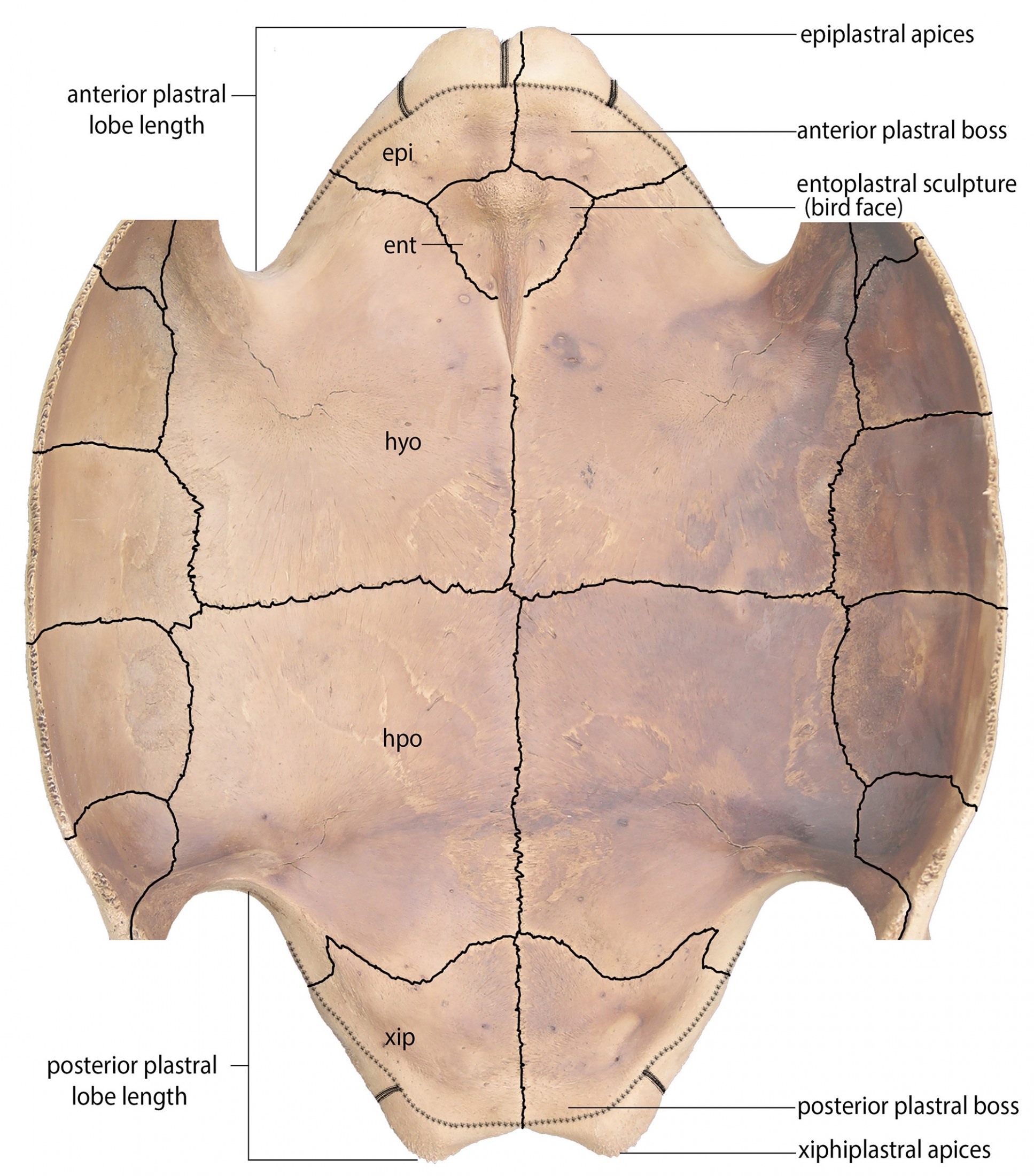
Epiplastral (Gular) Shelf.—In most tortoise genera, the epiplastral shelf forms a horizontal platform that extends from the gular projection to approximately the anterior suture of the entoplastron. The epiplastral shelf terminates as an abrupt elevated edge, often undercut, to form a shallow cavity for muscle attachment. The epiplastral shelf is prominent in mainland and Greater Antillean species of Chelonoidis, but absent from C. alburyorum. Preliminary observations suggest a similar reduction of the epiplastral shelf in Galápagos tortoises, although this needs further substantiation.
Plastral Boss.—Bosses are raised ornamentations or protuberances. In the thin-shelled C. alburyorum, boss-like structures occur as horizontal bony struts across midline sutures. If present, they may be located behind the epiplastral apices and/or just in front of the xiphiplastral apices. We suspect that these bosses help to reinforce vulnerable regions of the plastron in thin-shelled tortoises.
Entoplastral Sculptures.—Internal sculptures on the entoplastron function as places where the pectoral girdle and associated limb bones can exert force to allow front leg movements and assure assisted power for specialized behaviors, i.e., combat, navigating in broken terrain, and digging burrows. These sculptures typically have the appearance of a “bird-face” with a brow-line, a pair of “eye-socket” fossa, and a broad, elongated midline “beak.” The fragile distal end of this beak or keel is free from underlying plastral bone, and often is broken off in fossils. Features of the “bird-face” sculpture are consistent within species or subspecies, but show considerable variation among species and genera.
Position of Skin Lines.—As the sites of skin attachment to the shell, the skin lines are thin ridges, edges, or slight grooves on raised margins of the shell. The distance between the skin lines and the edges of the shell varies among Lucayan fossil populations. This variation is particularly evident on the epiplastron and the terminal edge of the xiphiplastron.
Plastral Lobe Sizes.—The anterior lobe (epiplastron + entoplastron + anterior part of hyoplastron) and the posterior lobe (xiphiplastron + posterior part of hypoplastron) show variation among species of Chelonoidis, particularly when comparing length versus width. Margins of the posterior lobe may be prominent, straight, or curvilinear. These differences may be sexually dimorphic in some populations.
First Dorsal Vertebra and Associated Ribs.— Features of the first dorsal (thoracic) vertebra and its two pairs of associated ribs, on the inside of the carapace, show variation, including differences in the shape and length of the centrum, presence or absence of the centrum keel, position of the anterior zygapophyses, and location of the ribs. Williams (1950b) recognized the usefulness of the first dorsal vertebra in describing the Mona Island fossil Chelonoidis (Monochelys) monensis as having an extremely elongated centrum, which he used to distinguish the subgenus Monochelys. Since then, others have employed the first dorsal vertebra in diagnoses, such as for Chelonoidis alburyorum (Franz and Franz, 2009), Chelonoidis dominicensis (Albury et al., 2018), and here in the new taxon from Grand Turk.
Rib Attachments.—Single ribs emerge from boney mounds on the internal surface of the costals; the free portions of these ribs extend upward to meet the accompanying vertebrae. The size, shape, and position of the rib mounds vary among populations, as do the length and shape of emerging free ribs; we suggest that together they offer possible characters.
Pelvic Articulation.—This apparatus consists of presacral and sacral ribs, their attachments to the inner wall of the carapace, and a massive articular surface. The final pair of presacral ribs arises from costal VII, followed by two pairs of sacral ribs that originate on costal VIII. A suture separates costal VII and VIII and their associated ribs. These three pairs of ribs rise from a thin, horizontal, web-like boney shelf, which includes the final pre-sacral ribs and two sacral ribs to form a broad articular surface that connects to the distal end of ilium. Robustness of the three pairs of ribs, the design of the bony platform that seats these ribs, and the size and orientation of the articular surface, all show variation and may have taxonomic value.
bones in black (upper; nuchal, neurals, pygal, and entoplastron not visible), and scutes in red (lower).
Abbreviations as in Figure 4.
morphological characters. Skin lines are indicated with a dotted line. Abbreviations as in Figure 4.
in black (left side) and scutes in red (right side). Posterior carapacial bones are identified in white letters.
Abbreviations as in Figure 4.
SYSTEMATIC PALEONTOLOGY
Order TESTUDINES
Family TESTUDINIDAE
Genus CHELONOIDIS
CHELONOIDIS ALBURYORUM ALBURYORUM Franz and Franz, 2009
Amended Diagnosis.—Prominent epiplastral (gular) shelf absent; plastron flattened (in presumed females only) or deeply concave (in presumed mature males only), each with a series of horizontal, boss-like boney ridges on the internal surfaces; small to indistinct epiplastral apices; moderately sized to nearly absent xiphiplastral apices; and presence of unique “bird-faced” entoplastral sculptures; extremely thin-shelled carapacial bones (see Figs. 4–8, 15 herein; also Franz and Franz, 2009:figs. 2–4, 11–12).
Remarks.—The lack of a prominent epiplastral (gular) shelf distinguishes Chelonoidis alburyorum from all species of Chelonoidis on Hispaniola, Cuba, and mainland South America. The Galápagos tortoises (Chelonoidis elephantopus sensu lato) also seem to lack the epiplastral shelf; their skeletal features are in need of greater morphological scrutiny. Differences in shell size, thickness and flatness of plastral bones, and degree of fusion of plastral sutures, suggest that sexual dimorphism occurred in some if not all populations of C. alburyorum.
Tortoise bones have been discovered on at least 14 Lucayan islands (Fig. 1). Among these populations, the only named taxon, until now, was Chelonoidis alburyorum Franz and Franz (2009) from Great Abaco on the Little Bahama Bank. Thus far, we have not evaluated in detail the morphology of the tortoise fossils from elsewhere in the island group, including Andros, Eleuthera, Long Island, Jumentos (Flamingo Cay), and New Providence (Great Bahama Bank), Crooked and Acklins (Crooked-Acklins Bank), San Salvador (San Salvador Bank), Mayaguana (Mayaguana Bank), or Great Inagua (Great Inagua Bank). While the available material seems to conform with the diagnostic characters of Chelonoidis alburyorum, much more work needs to be done to understand the tortoise radiation in the Lucayan Archipelago. Tortoises in general have considerable phenotypic plasticity (Fritz et al., 2005, 2007, 2010, 2012), so it seems reasonable that the populations from the Turks and Caicos be recognized as morphological variants of C. alburyorum.
In this paper, we will describe the tortoises from Middle Caicos (Caicos Bank) and Grand Turk (Turks Bank). The two new subspecies of Chelonoidis alburyorum that we are about to describe belong to the genus Chelonoidis because of these characters: flattened anterior plastron (i.e., no epiplastral shelf); and, cervical sulcus present on the nuchal instead of a cervical scute. We refer both new subspecies to C. alburyorum because of these characters: thin-walled carapace in both males and females; pelvic apparatus with one pair of presacral ribs on costal VII and two pairs of sacral ribs on costal VIII, terminating as an articular surface on suprapygal I, which connects to the pelvis; weakly developed epiplastral apices in both sexes; anterior plastral boss absent in males, weakly-developed in females; and, interior surface of entoplastron with a distinctive “bird-face” sculpture.
Chelonoidis alburyorum keegani
new subspecies
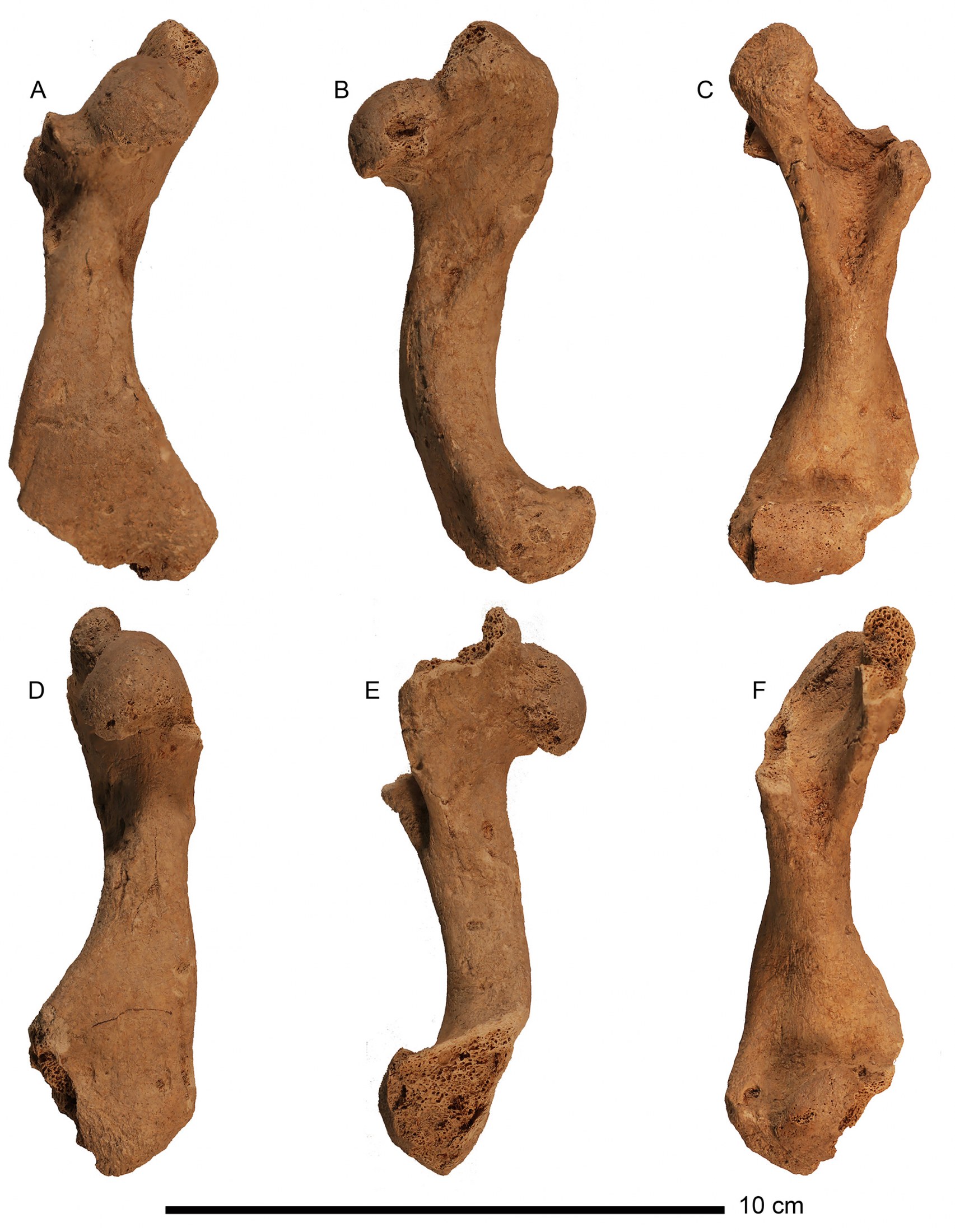
Holotype.—UF 453000, complete plastron, maturing male, reconstructed (Fig. 8A, C). Collected from FS 193, Unit 35, 96N, 100E, depth 41 cmbd, Coralie (site GT-3), Grand Turk, Turks and Caicos Islands.

(internal) aspect (A) and ventral (external) aspect (C), compared to the same views of C. a. alburyorum
(NMB.AB50.004; B, D, subadult). Note different scale bars.
Paratypes.—All from Coralie (site GT-3), Grand Turk, Turks and Caicos Islands. UF 452989, anterior plastron, maturing young male, partly reconstructed (Fig. 9A, C); from same provenience as holotype. UF 452991, anterior plastral lobe, mature female (Fig. 9B, D); from FS 348, Unit 75, 98N, 100E, 54–60 cmbd. UF 452990, partial plastron, much of it broken during excavation, large adult (old) male, anterior lobe partially reconstructed (Fig. 10); from FS 82, Unit 23, 37 cmbd. UF 452992, anterior plastral lobe, mature female (Fig. 11A, C); UF 452993, anterior plastral lobe, mature female (Fig. 11B, D). UF 452994, posterior plastral lobe, mature female, reconstructed (Fig. 12A, C). UF 452995, posterior plastral lobe, FRANZ ET AL.: Extinct tortoises from the Turks and Caicos Islands15 mature female, reconstructed (Fig. 12B, D). UF 452992–452995 all from FS 348, area 18, unit 5, 35–45 cmbd. TCNM.2020.001.01 posterior plastral lobe, maturing male (Fig. 13); from FS 236, Unit 47, 6.2–6.6 cmbd.

and ventral (external) aspect (C, D). A, C. UF 452989, maturing male. B, D. UF 452991, mature female.
Note different scale bars.

(internal) aspect (A), ventral (external) aspect (B), and in situ at the Coralie Archaeological site, Grand
Turk (C, where scale bar = 15 cm). Scale bar in upper right corner = 10 cm and applies to images A and B.

and ventral (external) aspect (C, D). A, C. UF 452992, mature female. B, D. UF 452993, mature female.
Note different scale bars.

and ventral (external) aspect (C, D). A, C. UF 452994, mature female. B, D. UF 452995, mature female

dorsal (internal) aspect (A) and ventral (external) aspect (B).
Type Locality.—Coralie Archaeological Site (GT-3). Latitude 21°30ʹ N, Longitude 71°08ʹ W, along the west shore of North Creek, on the northern end of Grand Turk Island, Turks and Caicos Islands, British Overseas Territory. Age is late Holocene (see above). Zoobank Nomenclatural Act.—22EDE7C913DC-46DC-9231-B91DFFD2053B.
Diagnosis.—Medium to large-sized tortoises, PL est. 345–600 mm, CL est. 431–710 mm, for subadult to adult males; PL est. 335–360 mm, CL est. 414–447 mm, for adult females (estimates based on comparisons with Chelonoidis a. alburyorum). Distinguished from C. a. alburyorum by: entoplastron “bird-face” with deep acromial fossae that accentuate a tall, slender, antero-posterior ridge; enlarged inguinal scutes extending from the posterior struts well onto the hypoplastron; xiphiplastral apices and anal notch prominent in males but weak to absent in females; posterior plastral boss well developed in both males and females; and centrum of first dorsal vertebra with a constricted waist, but lacking a prominent ventral keel.
Etymology.—We are pleased to name this extinct subspecies of tortoise in honor of William F. Keegan, Curator of Caribbean Archaeology, Florida Museum of Natural History, University of Florida, who initially saw the potential of the Coralie site. Keegan worked closely with Lizabeth A. Carlson in her dissertation research at Coralie. Their stellar efforts at the Coralie site, and at so many other Lucayan archaeological sites, made our research possible.
Specimen Provenience.—The tortoise skeletal remains from GT-3 were collected with numer- ous FS numbers. Two uncharred plastral, the holotype (UF 453000) and one paratype (UF 452989) were found at FS 193. They represent two male individuals. Five partial plastra (paratypes), carapace segments, and many isolated carapacial elements, all showed severe charring, derived from FS 348. All plastra in this sample were from females; we assume that the other exhumed elements from FS 348 were also from females, presumably asso- ciated with the plastra. Based on the numbers of plastra (3 anterior lobes, 2 posterior lobes), we suspect that the contents of this hearth represent three individual tortoises, perhaps from a single cooking event. Eleven specimens representing at least two individuals were recovered from FS 82. One of these was a large male, represented by UF 452990, a partial plastron. The other specimens from this feature are mostly limb elements from a much smaller individual (estimated CL 450 mm, based on associated humeri and carapaces from Sawmill Sink). The concentrated nature of the limb bones and presence of an intact plastron at FS 82 suggest the possibility of a kitchen midden, although the bones are not charred. Isolated tortoise remains were found scattered across the site at FS 41–42, 130, 147, 172, 219, 234, 235, 236, 274, 297, 340, and 343. These represents numerous individuals; some bones are incomplete; most are not charred. Their inclusion allows for greater morphological coverage of C. a. keegani, particularly with the elements not well represented in FS 82, 193, or 348.
Remarks.—The tortoise fossils from GT-3 belong to the genus Chelonoidis because of these characters: flattened anterior plastron (i.e., no epiplastral shelf); and cervical sulcus present on the nuchal instead of a cervical scute. We refer them to the species C. alburyorum because of these characters: thin-walled carapace in both males and females; pelvic apparatus with one pair of presacral ribs on costal VII and two pairs of sacral ribs on costal VIII, terminating as an articular surface on suprapygal I, which connects to the pelvis; weakly developed epiplastral apices in both sexes; anterior plastral boss absent in males, weakly-developed in females; and, interior surface of entoplastron with a distinctive “bird-face” sculpture (Franz and Franz, 2009).
Chelonoidis alburyorum sementis
new subspecies
Holotype.—UF 432441 (Fig. 14A–B), xiphiplastron, left, male. Collected from Unit 8, Indian Cave (site MC-37), Middle Caicos, Turks and Caicos Islands.
Paratypes.—All from Indian Cave (site MC-37). UF 432462 (Fig. 14C–D), epiplastron, left, Unit 7. UF 432442, epiplastron, Unit 8. UF 432455, epiplastron, Unit 5. UF 432493, epiplastron, Unit 8, 0–20 cm. UF 452983, hyoplastron, Unit 7, surface. UF 432444, hypoplastron/xiphiplastron/bridge peripheral VII, Unit 8. UF 432448, hypoplastron/bridge peripheral VII, Unit 7. UF 432467, hypoplastron with strut, Unit 7, surface. UF 452976, hypoplastron with strut, Unit 7. UF 452977, hypoplastron, Unit 7. UF 432488, xiphiplastron, Unit 7, surface. UF 432489, xiphiplastron, Unit 7, surface. UF 432490, xiphiplastron, Unit 7, surface. UF 432491, xiphiplastron, Unit 7, surface. UF 432469 (Fig. 14E–F), nuchal, left, Unit 7, surface.

Type Locality.—Indian (Head) Cave (site MC-37), 3 km west of the airport and Conch Bar Settlement, Middle Caicos Island, Turks and Caicos Islands, British Overseas Territory. Latitude: 21°49ʹ N, Longitude: 71°48ʹ W (Datum GWS 84). Age is late Holocene (see above).
Zoobank Nomenclatural Act.—63051E6B-78F2-4789-B526-792670231BF0.
Diagnosis.—Medium to large-sized tortoise, PL est. 345–480 mm, CL est. 450–600 mm for sub-adult to adult males (based on comparisons with complete shell of C. a. alburyorum from Franz and Franz, 2009; estimates for PL and CL of female C. a. sementis not available). Plastron of Chelonoidis a. sementis differs from that in C. a. keegani and C. a. alburyorum in having epiplastron with gular/ humeral sulcus less distinct, ventral (outer) surface more rugose, and posterior-lateral margin sharper (more dorso-ventrally compressed). Hypoplastron with lateral margin thinner but more rugose, and abdominal/femoral sulcus straighter (than in C. a. keegani only). Xiphiplastron thinner overall for its size, especially at lateral margin; in dorsal or ventral aspect, xiphiplastron much longer than it is wide (nearly square-shaped in the other two sub-species).
Etymology.—From the Latin sementis (feminine), meaning “a seeding, sowing” (Brown, 1956:693). The name Chelonoidis alburyorum sementis refers to the presumed ability of this extinct tortoise to disperse seeds of native plants in the Caicos Islands. The extinction of C. a. sementis and other tortoises in the Lucayan Archipelago resulted in the loss of keystone herbivores in these island ecosystems (Hastings et al., 2014).
Remarks.—Referral of the Indian Cave tortoise to C. alburyorum is based on the plastral characters listed above for C. a. keegani. The upper size limit in the diagnosis for C. a. sementis is based on an exceptionally large nuchal (UF 432469) from Unit 7. Other specimens of the subspecies are from smaller individuals.
DESCRIPTION OF CHELONOIDIS
ALBURYORUM KEEGANI
The type material of C. a. keegani consists of relatively intact plastra, which we describe here in detail. The other shell bones from GT-3 are isolated, typically incomplete specimens that we treat as referred specimens. The type series consists of three male and five female plastra; the males have thin plastral bones, loosely joined at the sutures. The only fully mature male in the series (UF 452990) has a deep plastral concavity that begins on the entoplastron; its plastron is nearly twice the size of that in the other, presumably younger males. All presumed female plastra (three anterior lobes plus two posterior lobes) are much more robust (thickened) with strongly fused sutures, weak apices, and essentially no xiphiplastral notch. We attribute these differences to sexual dimorphism.
Male Plastra
UF 453000 (Fig. 8A, C).—The holotype is a reconstructed, complete plastron from a maturing male; not associated with any axial, appendicular, or carapacial elements. The posterior lobe of the plastron has a shallow concavity, prominent apices, and a notch on the xiphiplastron, suggesting it has not reached full maturity. PL 343 mm; est. CL 431 mm, based on comparable measurements of C. a. alburyorum (Franz and Franz, 2009); PW 232 mm; other plastral measurements listed in Table 1.
No charring or butcher marks are present on the internal or external surfaces of UF 453000; it is pale grayish tan in color on both sides. Plastral bones are very thin-walled, and loosely connected to one another at the sutures by long splinter-like boney spears. Remnants of bridge peripherals attached to hyoplastron and hypoplastron; bridge peripherals enclosed by incomplete anterior and posterior bridge struts.
Interior surface lacks epiplastral shelf. Epiplastral apices weakly developed, having slightly rounded margins; a slight V-shaped indentation at the free margin where gular scutes meet the midline suture. Exterior surface of the entoplastron nearly rounded, slightly wider than long (68.6 x 58.1 mm); its interior surface similar in shape but smaller (precise measurements obscured by molding compound). Combined entoplastral and hyoplastral length 138 mm, or 40% of the length of the plastron, a calculation useful to estimate total length of the plastron. Xiphiplastron relatively short at midline, representing only about 40% of midline length of hypoplastron. Anal margin of the xiphiplastron arches inward to form a broad curvilinear notch, a condition found in males of living Chelonoidis carbonarius (Pingleton, 2009); notch terminates at each of the apices; skin line merges along the free edge of xiphiplastron near the notch.
Conspicuous raised skin line ridge along the anterior margin of internal shell edge, but no elevated boss on epiplastron; a horizontal boss crosses the midline in front of the xiphiplastral notch. Prominent entoplastral sculpture (“bird-face”) begins at anterior edge of the entoplastron, incorporating the entire element; sculpture with a heavy brow-line, moderately deep, paired eye-like fossae separated by a strong, prominent, beak-like keel; keel passes down the plastral midline, but broken as it nears the hyoplastron; the original beak extended beyond the entoplastron, terminating on hyoplastron.
Plastron with well-developed sulcal grooves that delineate the margins of each scute; gular scutes protrude slightly onto the entoplastron; the horizontally-positioned pectoral/abdominal sulci linear; pectoral scute 23.4 mm wide at midline, expanding outward as it approaches the bridge peripherals; anterior edge of the pectoral scute 15.7 mm below entoplastron; a small crescent-shaped axillary scute on anterior edge of frontal struts at the bridge; abdominal scute about 23% larger than femoral scute; the abdominal/femoral sulcus curvilinear arching anteriorly; and anal scutes rectangular, slanting forward to converge with the midline suture. One of the distinctive traits is the enlarged arrowhead-shaped inguinal scute, which crosses the bridge/hypoplastral suture and extends 57 mm onto the abdominal/femoral scutes, converging with the abdominal/femoral sulcus.
UF 452989 (Fig. 9A, C).—Partial plastron, maturing male, reconstructed. Specimen consists of right epiplastron (anterior edge eroded), entoplastron, partial right and left hyoplastra, with remnants of left and right anterior struts. PL est. 345 mm, CL est. 431 mm (based on plastral measurements from C. a. alburyorum); anterior lobe length 105 mm; PW est. 180 mm. Internal surface of plastron severely eroded; grayish tan in color, not charred. External surface fibrous, particularly in the strut region.
Sutures between the plastral bones deeply serrated and only weakly joined, as in holotype; sulci well defined. Right epiplastral apex with rounded margin, thickness 17.1 mm; hyoplastron thickness 5.2-5.9 mm. Skin line ridge narrow and steeply angled, along edge of epiplastron. Right gular scute short, intruding slightly onto entoplastron, which is oval in shape, with external surface wider (68.5 mm) than long (46.1 mm); its internal surface smaller (55.7 × 45.9 mm). “Bird-face” partially eroded on right side; the “eye socket” fossa moderately deep, brow-line moderately developed, and prominent beak broken where it separates from the underlying entoplastron near entoplastron/hyoplastron suture.
| MEASUREMENT | UF 453000 Young Adult Male |
UF 452990 Old Adult Male |
UF 452989 Maturing Young Male |
| Standard carapace length | Est. 431 | Est. 710 | Est. 431 |
| Midline length of plastron | 343 | Est. 600 | Est. 345 |
| Greatest length of plastron | 358 | — | — |
| Minimum bridge length | 192 | — | — |
| Entoplastron plus hyoplastron length | 138 | — | 142.1 |
| Anterior plastral lobe length< | 72 | 140 | 70 |
| Anterior plastral lobe width | Est. 180 | Est. 276 | — |
| Posterior plastral lobe length | 65 | — | — |
| Posterior plastral lobe width | 195 | — | — |
| Combined lobe lengths | 137 | — | — |
| Width across gular margins | 68 | 115 | — |
| Width across epiplastron/hyoplastron suture | 114 | 182 | — |
| Width across hypoplastron/xiphiplastron suture | 148 | — | — |
| Width across femoral/anal sulcus | 101 | — | — |
| Epiplastron length at midline | 34.3 | Est. 40 | — |
| Entoplastron length at midline | 59.4 | Est. 63 | 58.1 |
| Entoplastron width | 69.9 | 99.3 | 68.6 |
| Hyoplastron length at midline | 80 | — | 93.8 |
| Hypoplastron length at midline | 94< | — | — |
| Xiphiplastron length at midline | 59 | — | — |
| Distance between the entoplastron & humeral/pectoral sulcus | 15.7 | Est. 45 | 20.8 |
| Shell thickness at right hyoplastron | 4 | 3.1 | 5.4 |
| Shell thickness at right hypoplastron | 4.4 | — | — |
| Midline length of gular scute | 38.9 | Est. 80 | — |
| Max midline length of humeral scute | 71 | Est. 135 | 77 |
| Max midline length of pectoral scute | 23.4 | — | 28.9 |
| Max midline length of femoral scute | 80 | — | — |
| Width inguinal scute (from peripheral suture) | 44 | — | — |
| Max width of inguinal scute onto hypoplastron | 57 | — | — |
| Max length of inguinal scute | 44 | — | — |
| Midline length of anal scute | 20 | — | — |
| Depth anal notch | 12 | — | — |
| Width anal notch | 62 | — | — |
| Anal apices length from skin line | 13 | — | — |
Length of pectoral scute 27 mm at mid- line, expanding outward toward the left and right bridges; humeral/pectoral sulcus oblique, angling laterally to meet the free edge of hyoplastron just in front of anterior bridge struts. Pectoral/abdominal sulcus incomplete, but appears to form a straight line across the width of plastron before joining the peripheral edges. Right axillary scute crescent- shaped, 17.7 mm at widest point.
UF 452990 (Fig. 10A–C).—Reconstructed, partial plastron from an old male. It includes nearly complete right and left epiplastra, an entoplastron, and partial left side of the hyoplastron with anterior bridge strut. PL 600 mm (measurement taken during excavation), CL est. 710 mm (based on plastral measurements from C. a. alburyorum). Specimen was badly damaged when excavated, only partially reconstructed. Found lying upside down, atop wedge-shaped beach-sand deposit at 37 cmbs, ~50 m west of the marsh edge of North Creek (Fig. 2). External surface with a fibrous and pitted tex- ture. The buff-colored plastron has a large, eroded but boldly featured anterior lobe. Plastral lobe 150 mm midline length with prominent male concavity beginning midway on the entoplastron and continuing onto the hyoplastron. Wall of the concavity ascends steeply into the body cavity space, at least 45 mm above the general plastral plane; shell thickness on ascending wall 2.6 mm.
In internal (dorsal) view, epiplastron thick- ened (up to 27 mm) with a slightly elevated skin line and gular margins swollen; gulars squared off with little or no indentation at midline suture. The large entoplastron distorted from flexure of the male convexity that begins midway along the bone’s long axis. External width of entoplastron 98 mm, internal surface width 80 mm. Internal sur- face of entoplastron with exaggerated “bird-face” sculpture (exceptionally heavy brow line and two very deep “eye-socket” fossa on either side of a strongly elevated “beak” keel); beak 20 mm above the internal entoplastral surface, but broken off where separated from underlying plastral bone at the entoplastron/hyoplastron suture.
Plastral sulci with shallow grooves on the external shell surface. Left gular nearly complete; right gular heavily eroded. Base of gular scutes, where they converge near or at the anterior entoplastral suture, missing. Left strut with prominent, slightly crescent-shaped axillary scute, 50 mm high, 30 mm wide.
TCNM.2020.001.01 (Fig.13).—Maturing male, partial posterior plastral lobe consisting of right hypoplastron with strut and both xiphiplastra; includes developing plastral concavity of male; bones thin with loosely connected sutures; anal notch present; anal scutes rectangular; posterior horizontal boss, enlarged inguinal scute, hypoplastral/xiphiplastral suture curvilinear (not straight). Posterior lobe length 102 mm; posterior lobe width at struts 115 mm (right side only); xiphiplastron midline length 60 mm; width between anal apices 70 mm; anal notch length at midline 6 mm.
Female Plastra
Anterior Lobes: The anterior lobes of the plastron are extremely thickened, ranging in depth from 9 to 16 mm (versus 2–5 mm in male plastra), with strongly fused sutures. They have no evidence of the plastral depression (concavity) characteristic of males. The lateral margins of epiplastra are rounded; the weak to nearly absent epiplastral apices are slightly swollen with weak midline indentations; skin line scars and anterior bosses on epiplastra are weakly developed. The external surface of the entoplastron is diamond-shaped, whereas the internal surface has a prominent “bird-face” sculpture (heavy brow line, a shallow depression at midline of the brow, two shallow “eye-socket” fossa, separated by a prominent beak-like keel). The “beak” nearly always is broken where it diverges from underlying plastral bone. The sutural joints, securely binding the entoplastron to adjoining anterior plastral bones, consist of densely packed, tubular-shaped bony spears. These joints are well exposed along the internal margins of the ento- plastron, and expand outward to meet the external surface, a condition most readily observed in UF 452991, thereby producing a smaller internal entoplastral surface and a much more expansive external surface.
UF 452991 (Fig. 9B, D; Table 2).—Mature female, anterior plastral lobe, including both epiplastra, complete entoplastron, and nearly complete right hyoplastron. Both bridge struts missing. Internal surface well preserved, but external surface badly eroded, exposing cancellous and charred bone. Strong sutural joints (binding together the anterior plastral bones) densely packed, tube-shaped boney structures, well exposed along medial margins of entoplastron; they expand outward to meet the external entoplastral surface, producing a smaller internal entoplastral surface; external surface of entoplastron diamond-shaped, connected securely to adjoining bones.
UF 452992 (Fig. 11A, C).—Mature female, reconstructed, anterior plastral lobe. Complete left and right epiplastra with slightly eroded anterior margins, complete entoplastron, and nearly complete left and right hyoplastra. Both anterior struts absent. Internal surface of bones tan, uncharred; external surface blackened from charring, exposing some cancelleous bone. Anterior portion of lobe tilts upward in profile. Lateral margins of lobe relatively straight, slightly rounding anteriorly toward the epiplastral midline; margins of epiplastron eroded. The epiplastron possesses a well-defined skinline and pre-brow boss; anterior margin of the gulars entire, without a midline indenture. The entoplastron has the typical “bird-face” sculpture. Sutures strongly fused, although most sutures and sulci remain discernible; solidly constructed and flat (lacks plastral concavity). Epiplastral margins rounded on internal surface; skin line narrow at margins; a prominent horizontal boss between skin line and brow of the entoplastral “bird-face,” which resembles that in other tortoises from Grand Turk (prominent brow line, “eye-like” fossa, and central keel).
UF 452993 (Fig. 11B, D).—Mature female, anterior plastral lobe, including both epiplastra, complete entoplastron, most of left and right hyoplastra. Internal surface well preserved, but exterior surface mostly charred black with some exposed cancellous bone. Anterior boss weakly developed. The sutures are strongly fused. Epiplastron thickened, its rounded margins without obvious gular apices; skin lines weakly developed, barely visible along the plastral margins; a prominent horizontal boss lies between the skin line and the brow of the “bird face.”
Posterior Lobes: These two specimens probably are associated with two of the three anterior lobes but cannot be matched because of missing intervening bones. Both posterior lobes have remnants of bridge struts that are angular in cross-section, forming pronounced ascending lateral keels on their distal surfaces. Apices of the xiphiplastron are sheared off in both individuals, but the remnant edges suggest that the notches and apices were only weakly developed. They lack the plastral concavity characteristic of males. All three specimens include prominent elevated bar-like bosses extending across eroded terminus of the xiphiplastron.
UF 452994 (Fig. 12A, C).—Mature female, reconstructed posterior lobe including partial left and right hypoplastra, and left and right xiphiplastra. External shell surface charred; some cortical bone layer stripped off, exposing underlying cancelleous bone; internal surface uncharred. Xiphiplastral notch and apices incomplete. Posterior lateral boss near anal margin present. Hypoplastra with strongly keeled right strut; left strut missing. Length of posterior lobe at midline (less notch area) 76 mm; total lobe length with notch est. 84 mm; width of left side est. 100 mm; combined width between struts est. 208 mm; plastral thickness 11–18 mm.
UF 452995 (Fig. 12B, D).—Mature female, reconstructed posterior lobe including partial left and right hypoplastra, complete right xiphiplastron, incomplete left xiphiplastron. External surface charred exposing cancellous bone; internal shell surface without charring. Sutures partially visible; sulci obscure. Left hypoplastron missing strut; right hypoplastron with partial strut with strongly keeled internal margin. Posterior margins of xiphiplastra eroded, compromising measurements; xiphiplastral notch and apices incomplete; posterior horizontal boss present. Length of lobe at midline (less notch area) est. 68 mm; total length est. 75 mm; lobe width (calculated without left strut) est. 210 mm; plastral thickness 9–20 mm.
Referred Specimens From the Plastron
UF 453019, FS 219. Epiplastron, left, eroded, includes epiplastral apex. Gray patina, no charring. Epiplastral length at midline 29.4 mm. Gular/ humeral sulcus length to entoplastral suture 27.6 mm; gular sulcus encroaching slightly onto entoplastron, epiplastral apices small, weakly rounded lobe with slight indentation at midline; apices thickness 15.6 mm.
| MEASUREMENT | UF 452991 | UF 452992 | UF 452993 | UF 452994 | UF 452995 |
| Plastron length | Est. 335 | Est. 360 | Est. 355 | — | — |
| Carapace length | Est. 414 | Est. 447 | Est. 431 | — | — |
| Plastron width at epiplastron/hyoplastron suture | 116.4 | 125.6 | 122.5 | — | — |
| Plastron width at mid-point | — | — | Est. 210 | — | — |
| Anterior lobe length at midline | Est. 80 | Est. 87 | Est. 85 | — | — |
| Anterior lobe width between struts | — | — | Est 192 | — | — |
| Posterior lobe length | — | — | — | — | Est. 68 |
| Posterior lobe width | — | — | — | — | 210 |
| Shell thickness at bridge | — | 13.5 | 16 | 10 | 9 |
| Shell thickness at hyoplastron | 13.5 | 12 | 23.5 | — | — |
| Shell thickness at hypoplastron | — | — | — | 12.8 | 10.1 |
| Shell thickness at gular skin line | 18.9 | 20.4 | 20.1 | — | — |
| Epiplastron (midline) length | 33.4 | 38.6 | 41.3 | — | — |
| Entoplastron (external) length | Est. 55 | 56.5 | 59.9 | — | — |
| Entoplastron (external) width | Est. 72 | 68 | 78 | — | — |
| Entoplastron (internal) length | 41.4 | 45.1 | 44.4 | — | — |
| Entoplastron (internal) width | 50.1 | 57.4 | 58.9 | — | — |
UF 453020, FS 147. Entoplastron, internal surface with “bird-face,” external surface eroded; thickened with wide serrated sutural margins; brow line broad, with shallow “eye-socket” fossa, beak broad and less pronounced, extending to hyoplastron, where broken. Width est. 55.0 mm, midline length 50.1 mm.
UF 453021, FS 172. Plastron, posterior lobe, left. Length between posterior strut and xiphiplastral apices 82 mm.
UF 453027, FS 172. Hypoplastron/bridge peripheral, left, partial, texture rugose to fibrous. Bridge with enlarged partial inguinal scute extending onto abdominal scute, showing slight male concavity.
Referred Specimens From the Carapace
The carapacial elements from hearth FS 348 represent one to three individuals. The sample includes small pieces that we are unable to reconstruct. Bones of the prominently domed carapace(s) are very thin; posterior peripherals are nearly vertical in profile with slight flaring along their free margins; bridge peripherals and costals have similar vertical orientations. One nuchal has a prominent midline cervical sulcus, typical of the genus Chelonoidis; the other is too charred to evaluate; both specimens have a slight marginal indentation where the cervical sulci meet. The bridge peripherals also have weakly developed lateral keels. External surface of costals and peripherals lack growth annuli. The limited number of neurals and their condition prevent describing the complete neural formula, although those available have 4-sides (Neural I), 6-sides (Neural VII), and 4-sides (Neural VIII).
UF 452996, FS 348. Reconstructed, com- plete anterior carapacial rim (in two sections) from medium-sized tortoise, est. width 290 mm. External surfaces of nuchal and rim peripherals fire- scarred exposing cancelleous bone. Nuchal with cervical sulcus, lacking a cervical indentation; nuchal width 74 mm at marginal edge, 110.9 mm at broadest point.
UF 453004, FS 348. Partial nuchal with attached right peripheral I; neither surface charred; sutures strongly serrated. Free margin of peripheral width 59.2 mm, height 42 mm at posterior suture, height at sulcus 38.3 mm.
UF 453005, FS 348. Partial neural, missing anterior half, probably neural I, with remnant of vertebral spine; appears to be four-sided. External surface with faint horizontal sulcus, next to and paralleling the broken anterior part of neural; sutures strongly serrated; width at sulcus 45 mm; thickness along lateral margins 5.1 mm.
UF 453007, FS 348 (Fig. 15D). Reconstructed posterior carapace, both sides. Neurals VII-VIII (6- and 4-sided, respectively), costals VII-VIII, suprapygal I, part of suprapygal II. Internal surfaces with one pair of presacral ribs on costals VII; two pair of sacral ribs obliquely oriented, distally facing articulating surfaces on costals VIII.
UF 453006, FS 348. Two neurals, reconstructed, external surface charred; thickness adjacent to neural spine 5.3 mm. UF 453028, FS 41-42. Neural, 4-sided with lateral sulcus; strong vertebral spinal ridges; sulcus width 38.6 mm.
UF 453029, FS 41-42. Neural, 4-sided with lateral sulcus; strong vertebral spinal ridges; sulcus width 38.6 mm.
UF 452998, FS 348. Reconstructed, three joined left costals; middle costal (either III or V) wedge-shaped; maximum width 56 mm near neu rals, narrowing to 17.6 mm at junction of peripheral suture; thickness 4.7 mm; two adjoining costals incomplete. Two pieces removed for radiocarbon dating.
UF 452999, FS 348. Reconstructed, three joined costals (III-V) showing two partial vertebral scutes and two adjacent pleural scutes; sulci slightly raised; sutures moderately fused; thickness 3.0 and 3.2 mm.
UF 453008, FS 348. Partial left costals VII-VIII, with pelvic presacral, sacral ribs, and massive articulation. External surface charred. Right side, articulating buttress fragment.
UF 453026, FS 343. Peripheral I, right, incomplete; free edge of peripheral width 64.8 mm; skin line depth at sulcus 22.2 mm. Fragment removed for radiocarbon dating.
UF 453001, FS 348. Reconstructed, two joined right peripherals (II, III) at strongly serrated at costal suture, widths 43.1 and 44.9 mm. Free shell margin, entire, peripheral widths 51.8 and 41.8 mm; height of suture separating peripherals 54.4 mm; peripherals with vertical profile, flaring slightly outward at their free margins.
UF 452997, FS 348. Reconstructed, partial left carapace, composed of three bridge peripherals (IV-VI). Single weak horizontal keel across all three peripherals, just above the plastral-carapacial sutures. Width of peripheral V, 55.2 mm; thickness 3.1 mm.
UF 453018, FS 235. Peripheral V, bridge, incomplete. Lower surface rugose below marginal sulcus; sulcus between marginals V and VI on peripheral V with triangular notch midway along sulcus directed forward, a feature possibly unique to the genus Chelonoidis; thickness 3.1 mm, marginal sulcus width 62.8 mm.
UF 453002, FS 348. Reconstructed, left peripherals VII-VIII, including remnant of posterior strut; junction with costal sutures serrated; thickness along vertical sutures 4.7–5.8 mm.
UF 453030, FS 130. Peripheral XI, right, with peripheral/pygal suture; free margin slightly dentate at sulcus terminus; peripheral/costal sulcus width 51 mm; free margin edge width 78.5 mm; sulcus height 60.7 mm; thickness at midpoint 6.1 mm; skin line depth at sulcus 12.0 mm.
Referred Specimens From Axial Skeleton
UF 453002, FS 348 (Fig. 15A). Complete first dorsal vertebra; dorsal spine robust, firmly attached to neural I; anterior zygapophyses prominent, downward oriented, articular surfaces widely separated, forming a broad arch in dorso-frontal view; posterior zygapophysis weakly developed, barely discernible from its enlarged posterior base; total length 33.7 mm, maximum height above the shell 23.8 mm; centrum length 24.8 mm, with constricted waist 8.6 mm wide, no ventral keel; outside width between articular surfaces 29 mm, internal width 18 mm; neural canal rounded, width 4.1 mm.
UF 453003, FS 348 (Fig. 15B, upper image). Complete first dorsal vertebra, slightly more robust than UF 453002 (previous); neural bone absent; posterior zygapophysis weakly developed. Total length 32.8 mm, maximum height above the shell 25.1 mm; centrum length 22.3 mm, constricted waist width 9.5 mm, no ventral keel; anterior zygapophyses as in UF 453002, except for greater outside width between the zygapophyses articular surfaces 32 mm, internal width 17.4 mm, and with deeper arch. Neural canal oval in shape, height 7.4 mm, width 4.7 mm. UF 453003 is perhaps associated with UF 4543005, FS 348 (Fig. 15B, lower image), a partial neural I with fused 2nd dorsal vertebra.
UF 453022, FS 297. Large anterior caudal vertebra, eroded.
UF 453023, FS 297. Large anterior caudal vertebra, eroded.
UF 453024, FS 82. Mandibles, left and right, partial, from a small tortoise. Three pieces, same individual, external cutting edge serrated followed by a prominent groove.
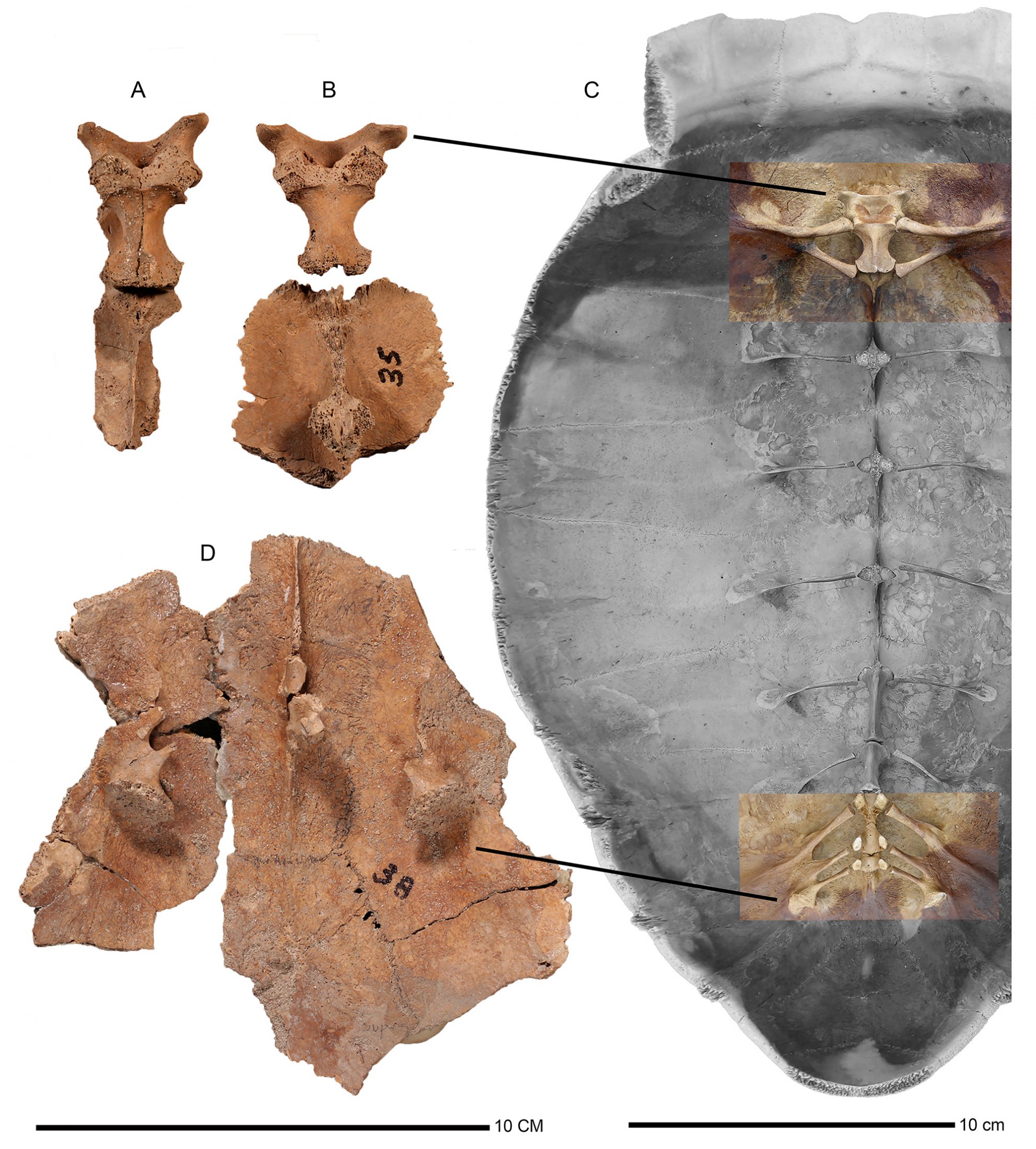
articulation (D). A. Chelonoidis alburyorum keegani (UF 453002). B. C. a. keegani (UF 453003; upper,
and UF 453005, possibly associated with UF 453003, lower). C. C. a. alburyorum (NMB.AB50.004). D.
Pelvic articulation of Chelonoidis alburyorum keegani (UF 453007). Note different scale bars.
Referred Specimens From the Appendicular Skeleton
UF 453009, FS 82. Scapula, right, missing coracoid except for a short “bud;” length 85.4 mm, acromion process length 46 mm; angle of scapula with respect to acromion process 131 degrees; glenoid fossa deeply arched with sutures meeting at mid-point; fossa length 26.1 mm x width 12.5 mm.
UF 453031, FS 274. Scapula, partial, with glenoid fossa; articular surface 20.6 x 10.8 mm.
UF 453010, FS 82 (Fig. 16A–C). Left humerus (paired with UF 453011), nearly complete except for right side of distal articulation; shaft recurved in profile; maximum length 95.9 mm, length of distal articulation to top of condyle 89.1 mm; condyle width 19.9 mm, length of distal articulation to intertrochanteric fossa 53.6 mm. Trochanteric tuberosities moderately divergent, separated by short, deep intertrochanteric fossa; greater trochanter extends 23 mm above crest of lesser trochanter; interior fossa width 17 mm between trochanters; fossa originates about half way up the shaft; shaft diameters 14.0 mm x 16.5 mm; M. latissimus dorsi scar shallow with roughened interior surface; distal external shaft groove shallow (but incomplete).
UF 453011, FS 82 (Fig. 16D–F). Right humerus (paired with UF 453010), greater and lesser trochanters broken, condyle nearly complete; shaft recurved in profile; total length est. 96 mm, length from distal articulation to top of condyle 92.4 mm, condyle width 20.1 mm, shaft diameters 14.1 x 16.9 mm, head 17 mm x 20 mm, slightly eroded on lower edge; external shaft groove shallow, complete length 21.6 mm, piercing through bone; M. latissimus dorsi scar shallow with roughened surface.
UF 453012, FS 82. Ulna, right, complete, eroded at distal end; total length 64.1 mm, shaft diameters 5.9 x 11.9 mm, width of articulation 18.5 mm, articular point 11.7 x 18.7 mm.
UF 453014, FS 82. Radius, matched pair; total length 55.4 mm (ends more eroded), midshaft diameters 7.2 mm x 7.2 mm.
UF 453015, FS 82. Femur, left, head and trochanters partially eroded; proximal fossa prominent; distal end missing; shaft diameters 15.5 mm x 13.2 mm.
UF 453025, FS 340. Femur, right, proximal and distal ends eroded.
UF 453016, FS 82. Tibia, left; total length 58.7 mm; proximal articulation width 20.3 mm, distal articulation width 14.8 mm; shaft flattened, diameters 8.1 mm x 11.2 mm.
UF 453017, FS 82. Fibula, left; total length 58.1 mm; shaft rounded, slender, diameters 6.0 mm x 7.4 mm.
aspect (B, E) and ventral aspect (C, F). A–C. UF 453010, left. D–F. UF 453011, right.
DESCRIPTION OF CHELONOIDIS
Alburyorum Sementis
The tortoise material from Indian Cave is from non-cultural contexts. The sample contains several hundred bones and bone fragments, although only about 100 of them are complete enough to provide useful information. None of these occurred as associated or articulated specimens from a single individual. Certain important elements are missing from the sample, which limits how thoroughly we can describe its morphology. Nevertheless, key plastral elements are present, which have characters that distinguish the Caicos tortoise from other subspecies of C. alburyorum. None of the plastral bones from MC-37 shows exceptionally thick shells or strong sutural fusions as in female plastra of the Grand Turk C. a. keegani. No entoplastron occurs among the specimens of C. a. sementis from MC-37.
Most plastral elements show features of males, with concavity development in five hypoplastral and four xiphiplastral specimens. Abrupt downward flexures of gular projections, initiated along the gular/humeral sulcus, are present in four epiplastral specimens. The anterior lobe is flattened without an epiplastral shelf. Gular projections are weakly developed, inflated, rounded, and with a shallow indentation at midline between the paired epiplastra. The anterior boss is absent. The gular/ humeral sulci converge at the anterior edge of entoplastral suture. Terminal margin of the xiphiplastral lobe is straight with apices and anal notch absent or weakly developed. The xiphiplastral boss is weakly defined. The anal scutes in xiphiplastra are rectangular.
Plastron (Holotype and Paratypes)
UF 432441 (Fig. 14A–B).—Xiphiplastron, left, male; thinner overall for its size, especially at lateral margin; in dorsal or ventral aspect, much longer than it is wide (nearly square-shaped in the other two species); margin without apex or notch; anal scute rectangular, well defined; flexed ventrally at femoral/anal sulcus with well-developed male concavity; moderately developed posterior boss; femoral/anal sulcus width 62.6 mm.
UF 432462 (Fig. 14C–D).—Epiplastron, left; gular/humeral sulcus oblique, flexed ventrally, length 31.1 mm; gular inflated, lateral margin rounded with V-shaped indentation at midline suture; gular scute meets entoplastral suture; anterior boss absent; midline suture length 24.9 mm.
UF 432455.—Epiplastron with gular/ humeral sulcus less distinct, ventral (outer) surface more rugose, and posterior-lateral margin sharper (more dorso-ventrally compressed).
UF 432467.—Hypoplastron with lateral margin thinner but more rugose, and abdominal/femoral sulcus straighter than in C. a. keegani.
Referred Specimens From the Plastron
UF 432442, Unit 8. Right epiplastron, badly weathered; lateral margin acute; flexed ventrally at gular/humeral sulcus; anterior boss absent; midline length 27.5 mm.
UF 432455, Unit 5. Left epiplastron; gular with raised skin line ridge on internal surface, lateral margin of epiplastron acute, slight indentation at midline suture; gular/humeral sulcus barely visible, oblique, flexed ventrally, length 31 mm; anterior boss absent; midline length 33.7 mm.
UF 432493, Unit 8, 0–20 cm. Right epiplastron, weathered, epiplastral apices eroded; gular with raised skin line ridge on internal surface, lateral margin of epiplastron acute, slight indentation at midline suture; gular/humeral sulcus oblique, flexed ventrally, length 33.7 mm; gular scute meets entoplastral suture; anterior boss absent; midline suture length 33.9 mm.
UF 452983, Unit 7, surface. Fragment of hyoplastron, left; sutures serrated; thickness 5.8 mm.
UF 432444, Unit 8. Hypoplastron/xiphiplastron/bridge peripheral VII, left partial, with strut, showing male concavity; with partial inguinal scute.
UF 432448, Unit 7. Hypoplastron/bridge peripheral VII, left, showing male concavity.
UF 432467, Unit 7, surface. Hypoplastron with strut, left, with male concavity.
UF 452976, Unit 7. Hypoplastron with strut, left, with male concavity.
UF 452977, Unit 7). Hypoplastron, right, large, partial, with partial inguinal scute.
UF 432488, Unit 7, surface. Left xiphiplastron; midline suture serrated; apex, notch, and posterior boss weakly defined; anal scute prominent; posterior margin width 32 mm, lateral margin 76 mm, femoral/anal sulcus 50.6 mm.
UF 432489. Unit 7, surface. Xiphiplastron, left, male; apex and notch weakly defined; lateral margin of anal scute acute; boss moderately enlarged; lateral margin length 58 mm.
UF 432490, Unit 7, surface. Xiphiplastron, left, immature male; midline suture 37 mm.
UF 432491, Unit 7, surface. Xiphiplastron, left; small individual; midline suture eroded; posterior margin straight, no apex or notch; skin line obscure; lateral margins acute; boss weakly defined.
Referred Specimens From the Carapac
Carapacial elements except peripherals are uncommon in the sample. A single partial nuchal has a midline cervical sulcus, which places this tortoise in the genus Chelonoidis. Two large costal pieces are very thin walled (<5 mm thick). Hundreds of very thin costal fragments, retrieved from cave sediments, perhaps were broken by foraging crabs. The 14 peripherals include six anterior, two bridge, two posterior, and four not evaluated for placement. The profiles and sizes of costals and peripherals suggest high-domed carapaces.
UF 432469 (Fig. 14E–F), Unit 7. Partial nuchal, right side only; bone broken across the cervical sulcus (incomplete), length est. 28 mm; total width est. 133 mm.
UF 432434, Unit 1, III/12. Neural I, complete, four-sided, vertebral spine attached; length 42 mm, width at sulcus 34 mm.
UF 452979, Unit 7. Partial costal I, with large rib attachment associated with first dorsal vertebra; shell thickness 3.1 and 4.7 mm.
UF 452980, Unit 7. Partial costal, large fragment with distal and proximal edges missing; lateral margins with sutures slightly serrated; sulcus groove very shallow; shell thickness 2.1 mm.
UF 432478, Unit 7. Peripheral I, right; length of free edge 84.5 mm, anterior suture line 50.2 mm, posterior suture line 54.9 mm, sulcus length 55.8 mm, maximum thickness at proximal suture 11.4 mm.
UF 432482, Unit 7. Peripheral I, right, partial; length of free margin est. 58 mm, maximum thickness 9.5 mm.
UF 432480, Unit 7. Peripheral I, left; length of free edge 71.5 mm, height of sulcus 48.3 mm, maximum thickness 12.7 mm.
UF 432483, Unit 7. Peripheral II, left; length of free margin 42.5 mm, height of sulcus 38.3 mm, maximum thickness 10.8 mm.
UF 432449, Unit 9. Bridge peripheral III fragment, left, with bridge strut, showing triangular-shaped inguinal scute.
UF 432453, Unit 9, 0–10 cmbs. Bridge peripheral VI, left; ventral portion with pitted texture; maximum thickness 4.7 mm.
UF 432456, Unit 7, surface. Bridge peripheral VII, left, partial, showing auxilliary scute.
UF 432477, Unit 7, surface. Peripheral VIII, right, free margin curvilinear, width 47 mm, height at anterior suture 57.5 mm, maximum thickness 5.0 mm.
UF 432484, Unit 7. Peripheral VIII, left; length of free margin 38.2 mm, height of sulcus 39.0 mm, maximum thickness 4.6 mm.
UF 432485, Unit 7. Peripheral IX, right; length of free margin 45.3 mm, height of sulcus 50.8 mm, maximum thickness 8.8 mm.
UF 432476, Unit 7, surface. Peripheral IX, left; length of free margin 43.9 mm, height at posterior suture 57 mm, height at anterior margin 64.6 mm, maximum thickness 9.5 mm.
UF 432481, Unit 7. Peripheral IX, right; length of free margin 49.9 mm, height of sulcus 51.4 mm, maximum thickness 11.2 mm.
UF 432452, Unit 9, 0–10 cmbs. Peripheral X, left; length of free margin 62.4 mm, sulcus height 51.1 mm, maximum thickness 4.3 mm.
UF 432479, Unit 7. Peripheral XI, near bridge, left; length of free margin 74 mm, anterior suture line 49 mm, height at sulcus 47.6 mm.
Referred Specimens From The Axial Skeleton
We recovered a considerable sample of cervical and caudal vertebrae, although they do not include the first dorsal vertebra.
UF 432458, Unit 7. Fragment of posterior cranium, includes full sagittal crest and foramen magnum.
UF 432468, Unit 7. Left mandible complete, right mandible partial; connected at symphysis; left oral groove length 31.1 mm; height 12.4 mm.
The sample consists of eight incomplete cervical vertebrae (UF 432457, 432459–432461, 452471, 452985–452987, probably representing at least three individuals. Most of the specimens are parts of the centrum, individual hypapophyses, and sections of the dorsal table. The four specimens described here demonstrate the general condition of the cervicals. Terminology follows Williams (1952).
UF 432457, Unit 7. Nearly complete; anterior articulation of centrum biconcave, suggesting Cervical VI; includes both pairs of zygapophyses, the anterior pair short and diminutive, the posterior pair robust; ventral surface of centrum flattened with thin keel total length 42 mm; vertebral foramen width 10.4 mm.
UF 432459, Unit 7. Highly incomplete except for posterior zygapophyses, which strongly turn downward, suggesting Cervical VIII, which articulates with first dorsal vertebra; vertebral foramen width 8.9 mm.
UF 432460, Unit 7. Nearly complete, long, slender; both articulations of centrum eroded, anterior and posterior zygapophyses present; total length 43.4 mm; vertebral foramen width 7.7 mm.
UF 452471, Unit 6, 0–10 cm. Incomplete in posterior portion, with anterior zygapophysis; anterior articulation of centrum with single concavity; ventral surface with prominent keel; vertebral foramen width 8.9 mm.
Ten caudal vertebrae (UF 432436, 432439, 432 445, 432446, 432450, 432492, 452973, 452978, 452984, 452988) are included from various units at MC-37. The caudal vertebrae are more complete and better preserved than the cervicals. It is nearly impossible to establish an orderly caudal sequence, although they obviously originated in different sections of the tail based on lengths of the transverse process. We choose a representative sample from several points along the tail to illustrate the general features of the sample, which represents more than one individual tortoise.
UF 452973, Unit 1, III/15. Complete caudal vertebra, originating from near base of tail; large individual; width across transverse processes 60 mm (left process incomplete); vertebral foramen 10 mm.
UF 452984, Unit 7. Complete caudal vertebra, probably from mid-section of tail; small individual; centrum length 21 mm; width across transverse processes 32.4 mm; vertebral foramen 5.7 mm.
UF 432445, Unit 3, I/1. Partial, caudal vertebra presumably originating from near tip of tail; probably a large individual; centrum length 22 mm; width across transverse processes est. 29 mm (right side incomplete); vertebral foramen tiny (<2 mm).
UF 432436 (Unit 1, III/16). Complete caudal vertebra, originating from near tip of tail; small individual; centrum length 14 mm; width across transverse processes 26 mm; vertebral foramen completely closed.
Referred Specimens From the Appendicular Skeleton
Among the one nearly complete and five partial pectoral girdle elements, most consist of glenoid fossa with remnants of scapulae and acromion processes; a single isolated coracoid and one attached to the nearly complete pectoral girdle are fan-shaped.
Pectoral Girdle.—UF 432470, Unit 7. Complete, except a missing outer edge of coracoid; scapula length 84.2 mm, acromion process length 47.6 mm, coracoid process length 35.8 mm, length and width of glenoid fossa 24.4 x 10.6 mm; acromion/scapula angle 136°.
UF 432471, Unit 7. Glenoid fossa; 36.5 x 20.3 mm.
UF 432472, Unit 7. Glenoid fossa; 23.7 x 10.8 mm.
UF 432473, Unit 7. Glenoid fossa, with partial scapula; 22.1 x 6.6 mm.
UF 432474, Unit 7. Coracoid, left, partial; est. length 50 mm. UF 432475, Unit 7. Coracoid, right, partial; eroded.
UF 452982, Unit 7. Glenoid fossa and partial scapula; fossa 26.4 x 11.8 mm.
Humerus.—UF 432437, Unit 2, II/11. Left, missing dorsal and proximal ends; trochanters incomplete; M. latissimus dorsi scar small, shallow, recurved in profile; shaft at midpoint 16.4 x 17.0 mm.
UF 432438, Unit 7. Left, partial trochanters and fossa; shaft diameters 27 x 19.7 mm; M. latissimus dorsi scar broad and shallow.
UF 432443, Unit 8. Right, missing distal articulation; head of humerus 15.7 x 13 mm; well-developed M. latissimus dorsi scar length 12.8 mm; proximal end of greater trochanter eroded; intertrochanteric fossa wide, shallow; distal groove well developed, deep; shaft diameters 10 x 9 mm.
UF 432465, Unit 7. Right, partial; proximal end eroded, distal end missing; partial distal groove present; intertrochanteric fossa wide; M. latissimus dorsi scar shallow, length 19 mm; shaft diameters 14.9 x 16.5 mm.
UF 432466, Unit 7. Left, distal end missing, proximal end eroded; M. latissimus dorsi scar shallow, weakly developed; shaft diameters 21 x 25 mm.
UF 452981, Unit 9, 0–10 cmbs. Left, proximal end eroded, distal end absent; M. latissimus dorsi scar prominent but shallow with elevated ridges on both sides; shaft diameters 20.2 x 16.8 mm.
Radius.—UF 432435, Unit 1, III/12. Right, complete; length 55.5 mm; proximal width 16.5 mm; shaft diameters 5.6 x 6.8 mm; distal width 16.5 mm.
UF 452972, Unit 1, III/15. Right, proximal end missing, arthritic; very large individual; shaft diameters 12.5 x 14.7 mm; distal width 32.0 mm.
Ulna.—UF 432486, Unit 7. Left, complete; total length 75 mm; proximal width 27 mm; shaft diameters at midpoint 7.5 x 13.3 mm.
UF 432487, Unit 7. Right, complete; total length 48.5 mm; proximal width 17.9 mm; shaft diameters at midpoint 4.5 x 8.3 mm.
Fused Radius and Ulna.—UF 452974, Unit 1, III/16. Distal half of both fused elements; distal width 40.8 mm.
Femur.—UF 432451, Unit 9. Right, complete, both trochanters and head slightly eroded; trochanteric fossa shallow; muscle scar weakly developed; total length 68 mm; distal articulation width 21.5 mm; shaft diameters 8 x 8 mm, angular in cross section.
UF 432454, Unit 9, 0–10 cm. Right, distal end eroded, trochanters missing, head complete; shaft diameters 11.8 x 11.9 mm, angular in cross section.
UF 432463, Unit 7. Right, complete, with minor erosion on posterior side of head and distal articulation; deep fossa between head and trochanters; total length 68.3 mm; distal articulation width est. 24.4 mm; length and width of head 20.5 x 16.8 mm.
UF 432464, Unit 7. Left, proximal end, large; trochanteric fossa deep with partially eroded head and trochanters.
Tibia.—UF 432440, Unit 2, IV/14. Left, complete, large; total length 97 mm; proximal width 37.6 mm; shaft diameters 29.2 x 15.9 mm.
UF 432447, Unit 8. Right, complete; total length 50.1 mm; proximal width 16.8 mm.
UF 452970, Unit 1, II/10. Left, complete; total length 53.6 mm; proximal width 26.4 mm; shaft diameters 8.9 x 7.7 mm.
UF 452975, Unit 7. Left, complete; shaft diameters 10.7 x 8.0 mm.
DISCUSSION
Colonization of West Indian Islands
Once ancestral Geochelone/Chelonoidis tortoises reached South America from Africa, they dispersed across the Neotropical landscape, reaching southern Panama by the early to middle Miocene (R. Franz, J. Bourque, pers. obs.). They differentiated into two lineages based on skeletal morphology: the carbonarius group that includes Chelonoidis carbonarius and C. denticulatus, and the chilensis group with C. chilensis and the Galápagos species (Auffenberg, 1971). These lineages were corroborated recently in a molecular phylogeny (Kehlmaier et al., 2017), which also placed the extinct Bahamian C. alburyorum as sister to the chilensis group. This arrangement would argue that a member (perhaps extinct?) of the chilensis group colonized the Bahamian region within the past million years.
We speculate that tortoises floated on ocean currents into the Lucayan and Greater Antillean islands from northern South America, following late stages of development of the Panamanian isthmus (late Miocene), an event that blocked entry of cold Pacific Ocean currents into the Equatorial Atlantic basin and led to new circulation patterns in the Caribbean/Western Atlantic basin (Butzin et al., 2011). Two possible dispersal routes for tortoises seem worth considering, one associated with the Antilles Current (Rowe et al., 2013), and the other with the Caribbean/Yucatan currents (Gyory et al., 2013). These currents originate with the Equatorial Atlantic Ocean off the northeastern coast of South America, and either could have transported tortoises from South America into the Caribbean and Bahamian basins.
At least two, possibly as many as four or five, dispersal events could account for the distinctive tortoise assemblages that once occupied islands of the Lucayan Archipelago, Cuba, Hispaniola, Navassa Island, Sombrero Island, and Mona Island. We note here the lack of tortoise fossils from Jamaica, Cayman Islands, Puerto Rico, the Haitian side of Hispaniola, and most of the Lesser Antilles; these absences may not be sampling artifacts since these islands have been well explored by archaeologists and paleontologists over the past 70 years. Shorter dispersal events within the Lucayan islands may have involved both the use of direct, intra-bank land connections between islands that formerly had been separated during interglacial times of high sea levels, as well as over-water dispersal between banks.
Tortoises are well adapted for dispersing across stretches of open ocean. Their moderate to large body sizes and high-domed shells (Pritchard, 1979), thick skins, armoring with heavy scales, substantial lung capacities, and buoyant bodies (Patterson, 1973) make them effective ocean-going rafts. Some tortoises apparently can withstand months at sea, as demonstrated by an adult Aldabra tortoise (Dipsochelys dussumieri) that came ashore on an African beach, after weeks at sea (Gerlach et al., 2006).
Tortoises can exist without food and water for extended periods of time. Some tortoise species can hold weakly diluted urine in specialized bladders to supplement their water needs (Jorgensen, 1998). Captive C. carbonarius have been observed to immerse their entire heads as they drank, suggesting a possible nasal drinking adaptation (RF pers. obs.; Pingleton, 2009:78). Nasal drinking has been reported for Aldabra tortoises, a behavior enhanced by specialized nasal anatomy that increases the efficiency of water-uptake (Arnold, 1979; Gerlach, 2004). Nasal drinking could increase the effectiveness of procuring freshwater during oceanic travels.
Paleoecology
The modern climate of the Turks and Caicos Islands is arid tropical throughout the year; highest temperatures occur from May through October, with typical daytime highs from 32 to 34°C from July to September. Cooler temperatures occur from December to March. Annual rainfall averages 700–750 mm for Grand Turk, with most rain from October to December. The hurricane season extends from June to November.
No permanent freshwater sources occur on Great Turk (Carlson, 1999) or Middle Caicos; a few depressions, presented on maps as the North and South wells on Grand Turk, may hold freshwater seasonally (W.F. Keegan, pers. com.). Armstrong Pond and other depressional wetlands on Middle Caicos are all salt ponds. Tortoises living under these water-stressed circumstances presumably obtained water from their foods or opportunistic drinking during rain events. In time, a population may have acquired anatomical, physiological, and behavior adjustments that capitalized on water retention and drinking efficiency (see examples inAuffenberg, 1963; Gerlach, 2004; Hewitt, 1933; Medica et al., 1980).
Correll and Correll (1982) described nine distinctive plant communities in the Lucayan Archipelago. Tortoises probably occupied each of these communities with the possible exception of coastal halophytic-dominated habitats. Optimal tortoise habitats would offer sufficient herbaceous ground cover, edible fruiting species, cactus, micro-sites for thermoregulation and refuge, friable soils suitable for nesting, and surface temperature regimes for production of mixed sex progeny. Tortoises may have moved seasonally among habitats to accommodate their changing needs or to avoid adverse weather conditions.
Dry coppice (broadleaf forest) is the dominant original vegetation type in the TCI. Pineyards (pinelands), though native to the Caicos Islands, are unknown in the Turk Islands. Most pieces of wood charcoal recovered from GT-3 were from coppice and coastal species, including wild lime, palm, buttonwood, and black torch (Carlson, 1999). We suggest that coppice trees were important by providing tortoises with carbohydrate-rich foods, specifically large edible fruits (>8 mm) (Patterson, 2002). Lucayan tortoises probably fed extensively on fallen fruits, as demonstrated by the massive fruit-accumulations (from mastic and satin-leaf plants) in the gut-cavity of two fossil specimens of Chelonoidis alburyorum from Sawmill Sink in the northern Bahamas (Franz and Franz, 2009).
Prior to the arrival of humans, adult tortoises in the Lucayan islands had no predators other than Cuban crocodiles (Crocodylus rhombifer), which are common fossils in most of the archipelago (Morgan and Albury, 2013; Hastings et al., 2014), although they have not been found in the Turks & Caicos. Other predators could have taken tortoise eggs and hatchlings, but would have been ineffectual in harming maturing and adult tortoises. These potential predators included Cuban crows (Corvus nasicus), Bahamian red-shouldered hawks (Buteo quadratus), magnificent frigatebird (Fregata magnificens), large land crabs (Cardisoma guanhumi), and Caribbean hermit crab (Coenobita clypeatus), and perhaps others.
Acknowledgements
We wish to thank all field crews who worked on the excavations and mapping at the Coralie and Indian Cave sites. We wish to single out Betsy Carlson, Bill Keegan, Michael Lace, John and Joan Mylroie, Michael Pateman, Brian Riggs, Anne Stokes, and Barbara and the late Reid Toomey for their hard work and dedication to Lucayan prehistory. We thank Richard Hulbert and Verity Mathis for valuable assistance in the museum collections. NSF grant GSS-149614 to DWS supported this research. The manuscript was improved by comments from Uwe Fritz and an anonymous reviewer.
Literature Cited
Albury, N., R. Franz, R. Rimoli, P. Lehman, and A. L. Rosenberger. 2018. Fossil land tortoises (Testudines: Testudinidae) from the Dominican Republic, West Indies, with a description of a new species. American Museum Novitates 3904:1–28
Arnold, E. N. 1979. Indian Ocean giant tortoises: their systematics and island populations. Pp.127–145. in D. R. Stoddart and T. S. Westoll, eds. The Terrestrial Ecology of Aldabra. Philosophical Transactions, Royal Society of London, 286.
Auffenberg, W. 1963. A note on the drinking habits of some land tortoises. Animal Behavior 11(1): 72–73.
Auffenberg, W. 1967. Notes on West Indian tortoises. Herpetologica 23:34–44.
Auffenberg, W. 1971. A new fossil tortoise, with remarks on the origins of South American testudines. Copeia 1971(3):106–117.
Auffenberg, W. 1974. Checklist of fossil land tortoises (Testudinidae). Bulletin of Florida State Museum, 18:316–251.
Bramble, D. M. 1971. Functional morphology, evolution, and paleoecology of gopher tortoises. Ph.D. dissertation, University of California, Berkeley. 341 pp.
Brown, R. H. 1956. Composition of Scientific Words. Smithsonian Institution, Washington, D.C. 882 pp.
Butzin, M., G. Lohmann, and T. Bickert. 2011. Miocene ocean circulation inferred from marine carbon cycle modeling combined with benthic isotope records. Paleooceanography 26, PA1203, doi:10.1029/2009PA001901
Carlson, L. A. 1999. Aftermath of a feast: Human colonization of the southern Bahamian Archipelago and its effects on the indigenous fauna. Ph.D. dissertation, University of Florida, Gainesville.
Carlson, L. A., and W. F. Keegan. 2004. Resource depletion in the prehistoric northern West Indies. Pp. 85–107 in S. M. Fitzpatrick (ed) Voyages of Discovery: The Archaeology of Islands. Westport, CT: Praeger Publishers.
Carlson, L. A., D. W. Steadman, and W. F. Keegan. 2006. Birdland: extinction steadily reduced a major food in the Taino diet. Times of the Islands (Turks & Caicos Islands) no. 74:64–69.
Correll, D. S., and H. B. Correll. 1982. Flora of the Bahamian Archipelago. J. Cramer. A.R. Ganter Verlg KG., FL-9490 Vaduz, 1692 pp.
Franz, R. 1999. Giant tortoises of the Turks & Caicos. Times of the Islands (Turks & Caicos Islands) Spring 1999: page 48.
Franz, R., and C. A. Woods. 1983. A fossil tortoise from Hispaniola. Journal of Herpetology 17:79–81.
Franz, R., L. A. Carlson, R. Owen, and D. W. Steadman. 1999. Fossil tortoises from the Turks and Caicos Islands, BWI. Pp. 27–31. in Clark-Simpson and Smith, eds. Proceeding 8th Symposium on the Natural History of the Bahamas. Gerace Research Center, San Salvador, Bahamas.
Franz, R., and S. E. Franz. 2009. A new fossil land tortoise in the genus Cheloniodis (Testudines: Testudinidae) from the northern Bahamas, with an osteological assessment of other Neo-tropical tortoises. Florida Museum of Natural History Bulletin 49(1):1–44.
Fritz, U., P. Široký, H. Kami and M. Wink. 2005. Environmentally caused dwarfism or a valid species – Is Testudo weissingeri Bour, 1996 a distinct evolutionary lineage? New evidence from mitochondrial and nuclear genomic markers. Molecular Phylogenetics and Evolution 37:389–401.
Fritz, U., A. K. Hundsdörfer, P. Široký, M. Auer, H. Kami, J. Lehmann, L. F. Mazanaeva, O. Türkozan and M. Wink. 2007. Phenotypic plasticity leads to incongruence between morphology-based taxonomy and genetic differentiation in western Palaearctic tortoises (Testudo graeca complex; Testudines, Testudinidae). Amphibia-Reptilia 28:97–121.
Fritz, U., S. R. Daniels, M. D. Hofmeyr, J. González, C. L. Barrio-Amorós, P. Široký, A. K. Hundsdörfer and H. Stuckas. 2010. Mitochondrial phylogeography and subspecies of the wide-ranging sub-Saharan leopard tortoise Stigmochelys pardalis (Testudines: Testudinidae) – a case study for the pitfalls of pseudogenes and GenBank sequences. Journal of Zoological Systematics and Evolutionary Research 48:348–359.
Fritz, U., L. Alcalde, M. Vargas-Ramírez, E. V. Goode, D. U. Fabius-Turoblin and P. Praschag. 2012. Northern genetic richness and southern purity, but just one species in the Chelonoidis chilensis complex. Zoologica Scripta 41:220– 232.
Gerlach, J. 2004. The complex nasal structure of Dipsochelys and its identification as a true Jacobson’s organ. Herpetolological Journal 15:15–20.
Gerlach, J., C. Muir, and M. D. Richmond. 2006. The first substantiated case of trans‐oceanic tortoise dispersal. Journal of Natural History 40(41–43):2403–2408. doi.org/10.1080/ 00222930601058240
Gyory, J., A. J. Mariano, and E. H. Ryan. 2013. “The Caribbean Current” Ocean Surface Currents. Retreived from https://oceancurrents.rsmas.miami.edu/caribbean/caribbean.html.
Hastings, A. K., J. Krigbaum, D. W. Steadman, and N. A. Albury. 2014. Reptile-dominated terrestrial food web in the Bahamas prior to human occupation. Journal of Herpetology 48:380– 388.
Hewitt, J. 1933. On the cape species and subspecies of the genus Chersinella Gray. Part I. Annals Natal Museum7:255–293.
Hulbert Jr., R. C. (editor). 2001. The fossil vertebrates of Florida. University Press of Florida, Gainesville. 350 pp.
Jorgensen, C. B. 1998. Role of urinary and cloacal bladders in chelonian water economy: historical and comparative perspectives. Biological Reviews of Cambridge Philosophical Society 73(4):347–366. doi.10.1017/ S0006323198005210
Keegan, W. F. 1997. Bahamian Archaeology: Life in the Bahamas and Turks and Caicos before Columbus. Media Publishing, Nassau, Bahamas, 104 pp.
Kehlmaier, C., A. Barlow, A. Hastings, M. Vamberger, J. L. A. Paipmano, D. W. Steadman, N. Albury, R. Franz, M. Hofreiter, and U. Fritz. 2017. Tropical ancient DNA reveals relationships of the extinct Bahamiam giant tortoise Chelonoidis alburyorum. Proceedings of the Royal Society B284:2016–2235. doi:10.1098/rspb.2016.2235
Leidy, J. 1868. Notice of some vertebrate remans from the West Indian islands. Proceedings of the Academy of natural Sciences of Philadelphia 20:178–180.
Medica, P. A., R. B. Bury, and R. A. Luckenbach. 1980. Drinking and construction of water catchments by the desert tortoise, Gopherus agassizii, in the Mojave Desert. Herpetologica 36(4):301–304.
Morgan, G. S., and N. A. Albury. 2013. The Cuban crocodile (Crocodylus rhombifer) from late Quaternary fossil deposits in the Bahamas and Cayman Island. Bulletin of Florida Museum of Natural History 52:161–236.
Olson, S. L., and J. R. Humphrey. 2017. The island and origin of Richard Harlan’s Galapagos tortoise Testudo elephantopus. Archives of Natural History 44:110–117.
Oswald, J. A., and D. W. Steadman. 2018. The late Pleistocene bird community of New Providence, Bahamas. The Auk: Ornithological Advances 135:359–377. Doi: 10.1642/AUK-17-185.1
Patterson, J. 2002. Native trees of the Bahamas. Bahamas National Trust. Media Enterprises, Nassau, Bahamas. Second ed, 128 pp.
Patterson, R. 1973. Why tortoises float. Journal of Herpetology 7(4):373–375.
Pingleton, M. 2009. The Redfoot Manual, A Beginner’s Guide to the Redfoot Tortoise. Art Gecko Publishing, Champaign, Illinois. 156 pp.
Poulakakis, N. S., Glaberman, M. Russello, L. B. Beheregaray, C. Ciofi, J. R. Powell, and A. Caccone. 2008. Historical DNA analysis reveals living descendants of an extinct species of Galápagos tortoise. PNAS doi/10.1073/pnas.0805340105
Poulakakis N. S., D. L. Edwards, Y. Chiari, R. C. Garrick, M. A. Russello, E. Benavides, G .J. Watkins-Colwell, S. Glaberman, W. Tapia, J. P. Gibbs, L. J. Cayot, and A. Caccone. 2015. Description of a new Galapagos giant tortoise species (Chelonoidis ; Testudines: Testudinidae) from Cerro Fatal on Santa Cruz Island. PLoS One. 2015 Oct 21;10(10):e0138779. doi: 10.1371/journal.pone.0138779. PMID: 26488886; PMCID: PMC4619298.
Pritchard, P. C. H. 1979. Encyclopedia of Turtles. T. F. H. Publ., Inc LTD, Neptune, NJ. 895 pp.
Rowe, E., A. J. Mariano, and E. H. Ryan. 2013. “The Antilles Current.” Ocean Surface Currents. https://oceancurrents.rsmas.miami.edu/ atlantic/antilles.html.
Steadman, D. W. 1999. Extinct animals in the West Indies. Times of the Islands (Turks & Caicos Islands), Spring 1999: page 47.
Steadman, D. W., R. Franz, G. S. Morgan, N.Albury, S. E. Franz, B. Kakuk, K. Broad, K.Tinker, M. Pateman, T. Lott, D. Jarzen, and D. Dilcher. 2007. Exceptionally well preserved late Quaternary plant and vertebrate fossils from a blue hole on Abaco, The Bahamas. Proceedings of the National Academy of Sciences of the United States of America 104(50):19897–19902.
Steadman, D. W., and J. Franklin. 2015. Changes in a West Indian bird community since the late Pleistocene. Journal of Biogeography 42: 426–438.
Steadman, D. W., N. A. Albury, L. A. Carlson, R. Franz, M. J. LeFebvre, B. Kakuk, and W. F. Keegan. 2019. Chronology of tortoise extinc-ion in the Bahamian Archipelago. The Holocene DOI: 10.1177/0959683619887412
Turvey, S. T, J. Almonte, J. Hansford, R. P. Scofield, J. L. Brocca, and S. D. Chapman. 2017. A new species of extinct late Quaternary giant tortoise from Hispaniola. Zootaxa 4277(1):1– 16. doi.org/10.11646/zootaxa.4277.1.1
Williams, E. E. 1950a. Testudo cubensis and the evolution of Western Hemisphere tortoises. Bulletin of the American Museum of Natural History 95:1–36.
Williams, E. E. 1950b. Variation and selection in certain central articulations of living turtles. Bulletin of the American Museum of Natural History 94:509–561.
Williams, E. E. 1952. A new fossil tortoise from Mona Island, West Indies, and a tentative arrangement of the tortoises of the world. Bulletin of the American Museum of Natural History 99:541–560.